Co-reporter:Anna I. Erickson, Reta D. Sarsam, and Andrew J. Fisher
Biochemistry 2015 Volume 54(Issue 45) pp:6830-6841
Publication Date(Web):October 29, 2015
DOI:10.1021/acs.biochem.5b01000
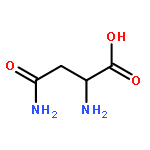
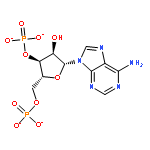
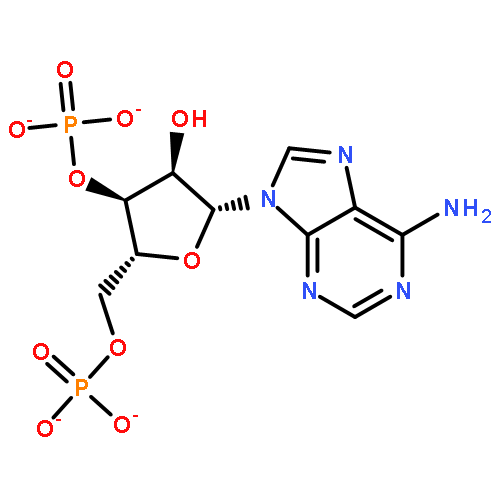
In many organisms, 3′-phosphoadenosine 5′-phosphate (PAP) is a product of two reactions in the sulfur activation pathway. The sulfurylation of biomolecules, catalyzed by sulfotransferases, uses 3′-phosphoadenosine 5′-phosphosulfate (PAPS) as a sulfate donor, producing the sulfated biomolecule and PAP product. Additionally, the first step in sulfate reduction for many bacteria and fungi reduces the sulfate moiety of PAPS, producing PAP and sulfite, which is subsequently reduced to sulfide. PAP is removed by the phosphatase activity of CysQ, a 3′,5′-bisphosphate nucleotidase, yielding AMP and phosphate. Because excess PAP alters the equilibrium of the sulfur pathway and inhibits sulfotransferases, PAP concentrations can affect the levels of sulfur-containing metabolites. Therefore, CysQ, a divalent cation metal-dependent phosphatase, is a major regulator of this pathway. CysQ (Rv2131c) from Mycobacterium tuberculosis (Mtb) was successfully expressed, purified, and crystallized in a variety of ligand-bound states. Here we report six crystal structures of Mtb CysQ, including a ligand-free structure, a lithium-inhibited state with substrate PAP bound, and a product-bound complex with AMP, phosphate, and three Mg2+ ions bound. Comparison of these structures together with homologues of the superfamily has provided insight into substrate specificity, metal coordination, and catalytic mechanism.
Co-reporter:Anna I. Erickson;Reta D. Sarsam
Acta Crystallographica Section F 2014 Volume 70( Issue 6) pp:750-753
Publication Date(Web):
DOI:10.1107/S2053230X14008619
CysQ is part of the sulfur-activation pathway that dephosphorylates 3′-phosphoadenosine 5′-monophosphate (PAP) to regenerate adenosine 5′-monophosphate (AMP) and free phosphate. PAP is the product of sulfate-transfer reactions from sulfotransferases that use the universal sulfate donor 3′-phosphoadenosine 5′-phosphosulfate (PAPS). In some organisms PAP is also the product of PAPS reductases that reduce sulfate from PAPS to sulfite. CysQ from Mycobacterium tuberculosis, which plays an important role in the biosynthesis of sulfated glycoconjugates, was successfully purified and crystallized in 24% PEG 1500, 20% glycerol. X-ray diffraction data were collected to 1.7 Å resolution using a synchrotron-radiation source. Crystals grew in the orthorhombic space group P212121, with unit-cell parameters a = 40.3, b = 57.9, c = 101.7 Å and with one monomer per asymmetric unit.
Co-reporter:Nhung Huynh, Aye Aye, Yanhong Li, Hai Yu, Hongzhi Cao, Vinod Kumar Tiwari, Don-Wook Shin, Xi Chen, and Andrew J. Fisher
Biochemistry 2013 Volume 52(Issue 47) pp:
Publication Date(Web):October 23, 2013
DOI:10.1021/bi4011754
N-Acetylneuraminate lyases (NALs) or sialic acid aldolases catalyze the reversible aldol cleavage of N-acetylneuraminic acid (Neu5Ac, the most common form of sialic acid) to form pyruvate and N-acetyl-d-mannosamine. Although equilibrium favors sialic acid cleavage, these enzymes can be used for high-yield chemoenzymatic synthesis of structurally diverse sialic acids in the presence of excess pyruvate. Engineering these enzymes to synthesize structurally modified natural sialic acids and their non-natural derivatives holds promise in creating novel therapeutic agents. Atomic-resolution structures of these enzymes will greatly assist in guiding mutagenic and modeling studies to engineer enzymes with altered substrate specificity. We report here the crystal structures of wild-type Pasteurella multocida N-acetylneuraminate lyase and its K164A mutant. Like other bacterial lyases, it assembles into a homotetramer with each monomer folding into a classic (β/α)8 TIM barrel. Two wild-type structures were determined, in the absence of substrates, and trapped in a Schiff base intermediate between Lys164 and pyruvate, respectively. Three structures of the K164A variant were determined: one in the absence of substrates and two binary complexes with N-acetylneuraminic acid (Neu5Ac) and N-glycolylneuraminic acid (Neu5Gc). Both sialic acids bind to the active site in the open-chain ketone form of the monosaccharide. The structures reveal that every hydroxyl group of the linear sugars makes hydrogen bond interactions with the enzyme, and the residues that determine specificity were identified. Additionally, the structures provide some clues for explaining the natural discrimination of sialic acid substrates between the P. multocida and Escherichia coli NALs.
Co-reporter:Nhung Huynh, Yanhong Li, Hai Yu, Shengshu Huang, ... Andrew J. Fisher
FEBS Letters (20 December 2014) Volume 588(Issue 24) pp:4720-4729
Publication Date(Web):20 December 2014
DOI:10.1016/j.febslet.2014.11.003
•Sialyltransferases couple sialic acid to glycans on glycoproteins and glycolipids.•Crystal structures of sialyltransferase from Photobacterium damselae have been determined.•Structure contains an N-terminal Ig-like domain with the GT-B sialyltransferase fold.•Binary structure with CMP-3F(a)Neu5Ac reveals a non-productive pre-Michaelis complex.Sialyltransferase structures fall into either GT-A or GT-B glycosyltransferase fold. Some sialyltransferases from the Photobacterium genus have been shown to contain an additional N-terminal immunoglobulin (Ig)-like domain. Photobacterium damselae α2–6-sialyltransferase has been used efficiently in enzymatic and chemoenzymatic synthesis of α2–6-linked sialosides. Here we report three crystal structures of this enzyme. Two structures with and without a donor substrate analog CMP-3F(a)Neu5Ac contain an immunoglobulin (Ig)-like domain and adopt the GT-B sialyltransferase fold. The binary structure reveals a non-productive pre-Michaelis complex, which are caused by crystal lattice contacts that prevent the large conformational changes. The third structure lacks the Ig-domain. Comparison of the three structures reveals small inherent flexibility between the two Rossmann-like domains of the GT-B fold.
Co-reporter:Burak V. Kabasakal, David D. Gae, Jie Li, J. Clark Lagarias, Patrice Koehl, Andrew J. Fisher
Archives of Biochemistry and Biophysics (15 September 2013) Volume 537(Issue 2) pp:233-242
Publication Date(Web):15 September 2013
DOI:10.1016/j.abb.2013.07.021
.jpg)
![Uridine 5'-(trihydrogendiphosphate), P'-[2-(acetylamino)-2-deoxy-a-D-glucopyranosyl] ester](http://img.cochemist.com/ccimg/600/528-04-1.png)
![Uridine 5'-(trihydrogendiphosphate), P'-[2-(acetylamino)-2-deoxy-a-D-glucopyranosyl] ester](http://img.cochemist.com/ccimg/600/528-04-1_b.png)
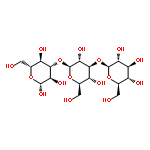






![5'-O-[hydroxy(sulfooxy)phosphoryl]adenosine 3'-(dihydrogen phosphate)](http://img.cochemist.com/ccimg/500/482-67-7.png)
![5'-O-[hydroxy(sulfooxy)phosphoryl]adenosine 3'-(dihydrogen phosphate)](http://img.cochemist.com/ccimg/500/482-67-7_b.png)








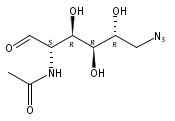
![D-Mannose, 2-[(2-azidoacetyl)amino]-2-deoxy-](/data/chemimg/113900/361154-23-6.png)
![D-Mannose, 2-[(2-azidoacetyl)amino]-2-deoxy-](/data/chemimg/113900/361154-23-6_b.png)
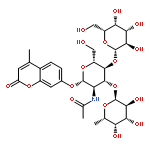
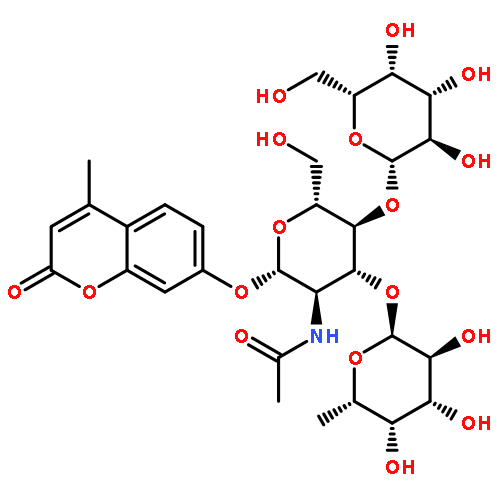

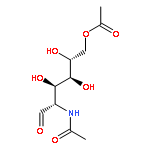
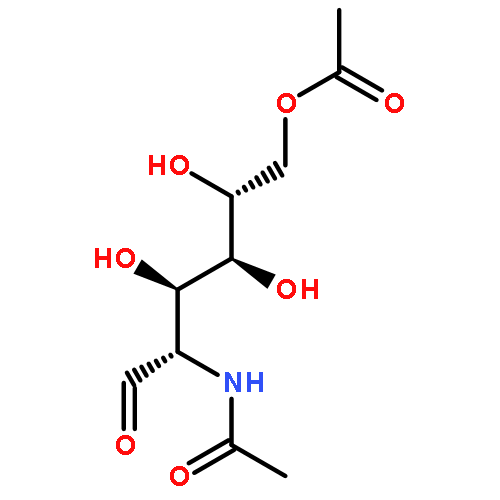
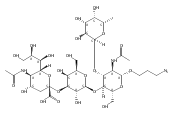
![2H-1-Benzopyran-2-one,7-[(4-O-b-D-galactopyranosyl-b-D-glucopyranosyl)oxy]-4-methyl-](http://img.cochemist.com/ccimg/84400/84325-23-5.png)
![2H-1-Benzopyran-2-one,7-[(4-O-b-D-galactopyranosyl-b-D-glucopyranosyl)oxy]-4-methyl-](http://img.cochemist.com/ccimg/84400/84325-23-5_b.png)
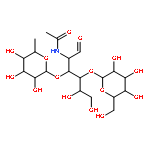
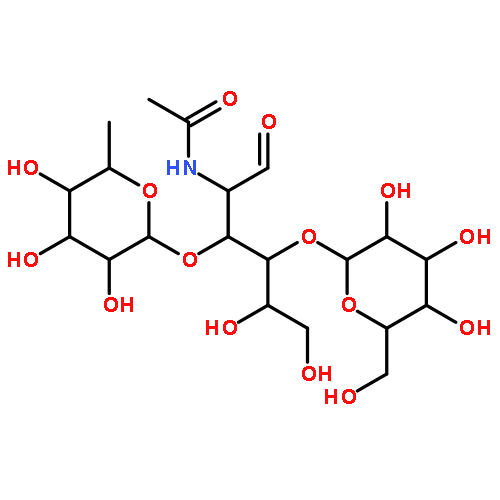






![D-Mannose, 2-deoxy-2-[(hydroxyacetyl)amino]-](http://img.cochemist.com/ccimg/7500/7483-19-4.png)
![D-Mannose, 2-deoxy-2-[(hydroxyacetyl)amino]-](http://img.cochemist.com/ccimg/7500/7483-19-4_b.png)