Co-reporter:Vidya Prasanna Kumar, Leonard M. Thomas, Kostyantyn D. Bobyk, Babak Andi, Paul F. Cook, and Ann H. West
Biochemistry 2012 Volume 51(Issue 4) pp:
Publication Date(Web):January 13, 2012
DOI:10.1021/bi201808u
Saccharopine dehydrogenase (SDH) catalyzes the final reaction in the α-aminoadipate pathway, the conversion of l-saccharopine to l-lysine (Lys) and α-ketoglutarate (α-kg) using NAD+ as an oxidant. The enzyme utilizes a general acid–base mechanism to conduct its reaction with a base proposed to accept a proton from the secondary amine of saccharopine in the oxidation step and a group proposed to activate water to hydrolyze the resulting imine. Crystal structures of an open apo form and a closed form of the enzyme with saccharopine and NADH bound have been determined at 2.0 and 2.2 Å resolution, respectively. In the ternary complex, a significant movement of domain I relative to domain II that closes the active site cleft between the two domains and brings H96 and K77 into the proximity of the substrate binding site is observed. The hydride transfer distance is 3.6 Å, and the side chains of H96 and K77 are properly positioned to act as acid–base catalysts. Preparation of the K77M and H96Q single-mutant and K77M/H96Q double-mutant enzymes provides data consistent with their role as the general acid–base catalysts in the SDH reaction. The side chain of K77 initially accepts a proton from the ε-amine of the substrate Lys and eventually donates it to the imino nitrogen as it is reduced to a secondary amine in the hydride transfer step, and H96 protonates the carbonyl oxygen as the carbinolamine is formed. The K77M, H976Q, and K77M/H96Q mutant enzymes give 145-, 28-, and 700-fold decreases in V/Et and >103-fold increases in V2/KLysEt and V2/Kα-kgEt (the double mutation gives >105-fold decreases in the second-order rate constants). In addition, the K77M mutant enzyme exhibits a primary deuterium kinetic isotope effect of 2.0 and an inverse solvent deuterium isotope effect of 0.77 on V2/KLys. A value of 2.0 was also observed for D(V2/KLys)D2O when the primary deuterium kinetic isotope effect was repeated in D2O, consistent with a rate-limiting hydride transfer step. A viscosity effect of 0.8 was observed on V2/KLys, indicating the solvent deuterium isotope effect resulted from stabilization of an enzyme form prior to hydride transfer. A small normal solvent isotope effect is observed on V, which decreases slightly when repeated with NADD, consistent with a contribution from product release to rate limitation. In addition, V2/KLysEt is pH-independent, which is consistent with the loss of an acid–base catalyst and perturbation of the pKa of the second catalytic group to a higher pH, likely a result of a change in the overall charge of the active site. The primary deuterium kinetic isotope effect for H96Q, measured in H2O or D2O, is within error equal to 1. A solvent deuterium isotope effect of 2.4 is observed with NADH or NADD as the dinucleotide substrate. Data suggest rate-limiting imine formation, consistent with the proposed role of H96 in protonating the leaving hydroxyl as the imine is formed. The pH–rate profile for V2/KLysEt exhibits the pKa for K77, perturbed to a value of ∼9, which must be unprotonated to accept a proton from the ε-amine of the substrate Lys so that it can act as a nucleophile. Overall, data are consistent with a role for K77 acting as the base that accepts a proton from the ε-amine of the substrate lysine prior to nucleophilic attack on the α-oxo group of α-ketoglutarate, and finally donating a proton to the imine nitrogen as it is reduced to give saccharopine. In addition, data indicate a role for H96 acting as a general acid–base catalyst in the formation of the imine between the ε-amine of lysine and the α-oxo group of α-ketoglutarate.
Co-reporter:Alla O. Kaserer, Babak Andi, Paul F. Cook and Ann H. West
Biochemistry 2009 Volume 48(Issue 33) pp:
Publication Date(Web):July 20, 2009
DOI:10.1021/bi900886g
The multistep His-Asp phosphorelay system in Saccharomyces cerevisiae allows cells to adapt to osmotic, oxidative, and other environmental stresses. The pathway consists of a hybrid histidine kinase SLN1, a histidine-containing phosphotransfer (HPt) protein YPD1, and two response regulator proteins, SSK1 and SKN7. Under nonosmotic stress conditions, the SLN1 sensor kinase is active, and phosphoryl groups are shuttled through YPD1 to SSK1, therefore maintaining the response regulator protein in a constitutively phosphorylated state. The cellular response to hyperosmotic stress involves rapid efflux of water and changes in intracellular ion and osmolyte concentration. In this study, we examined the individual and combined effects of NaCl and glycerol on phosphotransfer rates within the SLN1-YPD1-SSK1 phosphorelay. The results show that the combined effects of glycerol and NaCl on the phosphotransfer reaction rates are different from the individual effects of glycerol and NaCl. The combinatory effect is likely more representative of the in vivo changes that occur during hyperosmotic stress. In addition, the effect of osmolyte concentration on the half-life of the phosphorylated SSK1 receiver domain in the presence/absence of YPD1 was evaluated. Our findings demonstrate that increasing osmolyte concentrations negatively affect the YPD1·SSK1∼P interaction, thereby facilitating dephosphorylation of SSK1 and activating the HOG1 MAP kinase cascade. In contrast, at the highest osmolyte concentrations, reflective of the osmoadaptation phase of the signaling pathway, the kinetics of the phosphorelay favor production of SSK1∼P and inhibition of the HOG1 pathway.
Co-reporter:Ashwani K. Vashishtha, Ann H. West, Paul F. Cook
Archives of Biochemistry and Biophysics (15 October 2015) Volume 584() pp:
Publication Date(Web):15 October 2015
DOI:10.1016/j.abb.2015.08.023
•D126, C154, and Y99 were changed alone and in pairs to give mutant enzymes.•Small changes in rate constants suggest residues are not catalytically essential.•pH-rate profiles are qualitatively similar to those of wild type enzyme.•Data are consistent with the intramolecular mechanism proposed from QM/MM studies.•All residues mutated likely facilitate a conformational change to close the site.Saccharopine reductase catalyzes the reductive amination of l-α-aminoadipate-δ-semialdehyde with l-glutamate to give saccharopine. Two mechanisms have been proposed for the reductase, one that makes use of enzyme side chains as acid–base catalytic groups, and a second, in which the reaction is catalyzed by enzyme-bound reactants. Site-directed mutagenesis was used to change acid–base candidates in the active site of the reductase to eliminate their ionizable side chain. Thus, the D126A, C154S and Y99F and several double mutant enzymes were prepared. Kinetic parameters in the direction of glutamate formation exhibited modest decreases, inconsistent with the loss of an acid–base catalyst. The pH-rate profiles obtained with all mutant enzymes decrease at low and high pH, suggesting acid and base catalytic groups are still present in all enzymes. Solvent kinetic deuterium isotope effects are all larger than those observed for wild type enzyme, and approximately equal to one another, suggesting the slow step is the same as that of wild type enzyme, a conformational change to open the site and release products (in the direction of saccharopine formation). Overall, the acid–base chemistry is likely catalyzed by bound reactants, with the exception of deprotonation of the α-amine of glutamate, which likely requires an enzyme residue.
Co-reporter:Chaonan Hsu, Ann H. West, Paul F. Cook
Archives of Biochemistry and Biophysics (15 October 2015) Volume 584() pp:
Publication Date(Web):15 October 2015
DOI:10.1016/j.abb.2015.08.016
•Replacing D271 with N in homoisocitrate dehydrogenase gave 500-fold lower activity.•The same slow step limits at high and low homoisocitrate concentration.•The pH independence of V/Et indicates chemistry is not slow.•The effect of solvent deuterium suggests a slow pre-catalytic conformational change.Homoisocitrate dehydrogenase (HIcDH) catalyzes the NAD+-dependent oxidative decarboxylation of HIc to α-ketoadipate, the fourth step in the α-aminoadipate pathway responsible for the de novo synthesis of l-lysine in fungi. A mechanism has been proposed for the enzyme that makes use of a Lys–Tyr pair as acid-base catalysts, with Lys acting as a base to accept a proton from the α-hydroxyl of homoisocitrate, and Tyr acting as an acid to protonate the C3 of the enol of α-ketoadipate in the enolization reaction. Three conserved aspartate residues, D243, D267 and D271, coordinate Mg2+, which is also coordinated to the α-carboxylate and α-hydroxyl of homoisocitrate. On the basis of kinetic isotope effects, it was proposed that a conformational change to close the active site and organize the active site for catalysis contributed to rate limitation of the overall reaction of the Saccharomyces cerevisiae HIcDH (Lin, Y., Volkman, J., Nicholas, K. M., Yamamoto, T., Eguchi, T., Nimmo, S. L., West, A. H., and Cook, P. F. (2008) Biochemistry47, 4169–4180.). In order to test this hypothesis, site-directed mutagenesis was used to change D271, a metal ion ligand and binding determinant for MgHIc, to N. The mutant enzyme was characterized using initial rate studies. A decrease of 520-fold was observed in V and V/KMgHIc, suggesting the same step(s) limit the reaction at limiting and saturating MgHIc concentrations. Solvent kinetic deuterium isotope effects (SKIE) and viscosity effects are consistent with a rate-limiting pre-catalytic conformational change at saturating reactant concentrations. In addition, at limiting MgHIc, an inverse (SKIE) of 0.7 coupled to a significant normal effect of viscosogen (2.1) indicates equilibrium binding of MgHIc prior to the rate-limiting conformational change. The maximum rate exhibits a small partial change at high pH suggesting a pH-dependent conformational change, while V/KMgHIc exhibits the same partial change observed in V, and a decrease at low pH with a pKa of 6 reflecting the requirement for the unprotonated form of MgHIc to bind to enzyme. However, neither parameter reflects the pH dependence of the chemical reaction. This pH independence of the chemical reaction over the range 5.5–9.5 is consistent with the much slower conformational change that would effectively perturb the observed pK values for catalytic groups to lower and higher pH. In other words, the pH dependence of the chemical reaction will only be observed when chemistry becomes slower than the rate of the conformational change. Data support the hypothesis of the existence of a pre-catalytic conformational change coupled to the binding of MgHIc.
.jpg)

![[5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl [hydroxy-[(3,4,5-trihydroxyoxolan-2-yl)methoxy]phosphoryl] Hydrogen Phosphate](http://img.cochemist.com/ccimg/20800/20762-30-5.png)
![[5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl [hydroxy-[(3,4,5-trihydroxyoxolan-2-yl)methoxy]phosphoryl] Hydrogen Phosphate](http://img.cochemist.com/ccimg/20800/20762-30-5_b.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)












![(2R)-2-AMINO-1-PHENYL-PROPAN-1-OL; (E)-4-(4-BROMOPHENOXY)-4-OXO-B<WBR />UT-2-ENOIC ACID; N,N-DIMETHYL-3-(2-PYRIDYL)PROPAN-1-AMINE; 3-[(1R<WBR />)-1-HYDROXY-2-METHYLAMINO-ETHYL]PHENOL; DIHYDROCHLORIDE](http://img.cochemist.com/ccimg/400/320-77-4.png)
![(2R)-2-AMINO-1-PHENYL-PROPAN-1-OL; (E)-4-(4-BROMOPHENOXY)-4-OXO-B<WBR />UT-2-ENOIC ACID; N,N-DIMETHYL-3-(2-PYRIDYL)PROPAN-1-AMINE; 3-[(1R<WBR />)-1-HYDROXY-2-METHYLAMINO-ETHYL]PHENOL; DIHYDROCHLORIDE](http://img.cochemist.com/ccimg/400/320-77-4_b.png)













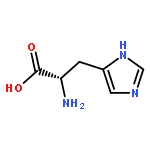
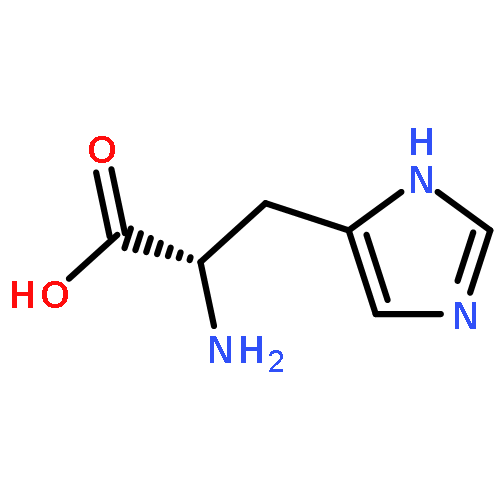
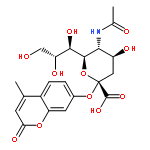
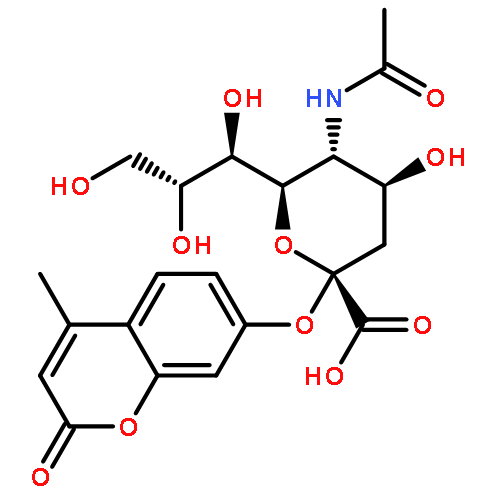
![(3AR,4R,5R,6AS)-4-FORMYL-2-OXOHEXAHYDRO-2H-CYCLOPENTA[B]FURAN-5-Y<WBR />L 4-BIPHENYLCARBOXYLATE](http://img.cochemist.com/ccimg/100/72-89-9.png)
![(3AR,4R,5R,6AS)-4-FORMYL-2-OXOHEXAHYDRO-2H-CYCLOPENTA[B]FURAN-5-Y<WBR />L 4-BIPHENYLCARBOXYLATE](http://img.cochemist.com/ccimg/100/72-89-9_b.png)
![L-Glutamic acid,N-[(5S)-5-amino-5-carboxypentyl]-](http://img.cochemist.com/ccimg/1000/997-68-2.png)
![L-Glutamic acid,N-[(5S)-5-amino-5-carboxypentyl]-](http://img.cochemist.com/ccimg/1000/997-68-2_b.png)