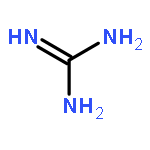
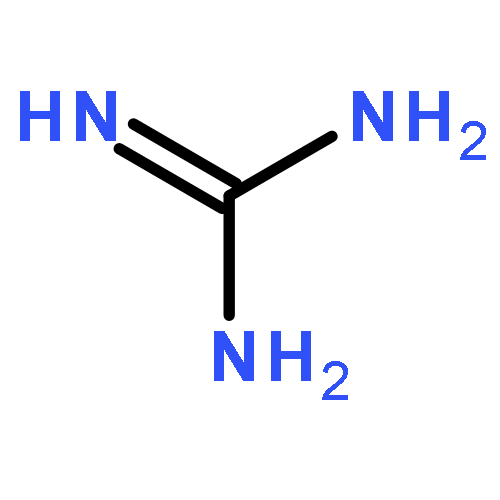
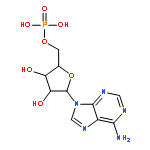
Co-reporter:Li-Dan Hu;Xiang-Jun Chen;Xiao-Yan Liao
Protein & Cell 2017 Volume 8( Issue 8) pp:618-622
Publication Date(Web):10 July 2017
DOI:10.1007/s13238-017-0430-6
Dear Editor,Modulation of mRNA transportation, localization, translational efficiency, and degradation plays an important role in the regulation of gene expression. In eukaryotic cells, translationally repressed mRNAs may be recruited into distinct intracellular foci termed as RNA granules, which are microscopically visible non-membrane-bound organelles composed of messenger ribonucleoproteins (mRNPs) (Anderson and Kedersha, 2009). Stress granule (SG) and processing body (P body) are two evolutionarily conserved cytoplasmic RNA granules in somatic cells (Anderson and Kedersha, 2006). Assembly of SGs can be stimulated by various stresses and SGs will be disassembled after the stress is removed. By shifting the equilibrium between stalled and translating mRNAs, SG formation can modulate stress response of the cells (Protter and Parker, 2016). It is increasingly recognized that aberrant SG assembly/disassembly may affect cell survival and human diseases thereafter (Anderson et al., 2015; Li et al., 2013; Mahboubi and Stochaj, 2017).SG assembly and disassembly can be influenced by many factors including various endogenous and extracellular stressors, eIF2α phosphorylation and overexpression of SG promoting proteins (Mahboubi and Stochaj, 2017; Panas et al., 2016; Protter and Parker, 2016). Particularly, several RNA-binding proteins such as TIA-1, TIAR, G3BP, CPEB1, and TTP can initiate SG formation and therefore overexpression of these proteins will promote SG formation even under non-stressed conditions (Kedersha et al., 1999; Stoecklin et al., 2004; Tourriere et al., 2003; Wilczynska et al., 2005). Chemical compounds can also modulate SG formation by affecting translation, proteasome activity or endogenous stressors (Mahboubi and Stochaj, 2017). It is worth noting that the chemical compounds identified thus far affect SG assembly/disassembly indirectly. Meanwhile, the action of these compounds may introduce complicated cellular responses and thereby it is difficult to study the role of SGs in specific cellular events. It has been reported that formation of several cellular bodies including SG, P body, and aggresome involves interactions between low-complexity sequences (Kato et al., 2012; Patel et al., 2015; Reijns et al., 2008). This suggests that these microscopic visible membraneless organelles may share some general rules in assembly though they have dissimilar components and morphology. An interesting question is whether there exist specific SG modulators. To address this problem, we screened a compound library obtained from Chinese traditional medical plants by a novel screening strategy (Fig. 1A).Open image in new windowFigure 1Screening SG-specific modulators using isolated SG cores from a Chinese traditional plant-derived compound library containing 116 compounds. (A) A schematic strategy for screening SG-specific chemical compounds. SG cores isolated from cells overexpressing GFP-TIA-1 were divided into small aliquots and treated with compounds from the library. Western blot analysis was used to determine the SG modulators. (B) Quantitative results of the effects of the 80 μmol/L compounds on SG assembly/disassembly in vitro. The data were calculated from the ratio of GFP-TIA-1 in the precipitation fraction to that in the supernatant fraction and normalized by taking the control data as 100%. The data of two highly nontoxic compounds are highlighted in red and green and the structures of these two compounds are also shown. The data are presented as mean ± S.E.M. calculated from three independent experiments. (C) Effects of benzene derivatives on SG stability. The structures show four highest activity compounds. (D) Effects of selected compounds with high activity on cell viability. The data are shown as mean ± S.E.M. (n = 3). *P < 0.05. (E) Upper panel: representative Western blot analysis of effects of C13 and C85 on isolated SG cores detected by the amounts of GFP-TIA-1 probed by GFP antibody. P and S are the precipitation and supernatant fraction, respectively. Lower panel: representative confocal microscope images of isolated SGs treated with 0.8% DMSO, C13, and C85 in 0.8% DMSO. Scare bar, 20 μm. (F) Size distributions of isolated SGs. The number of small (0.4–2 μm), medium (2–5 μm), and large SGs (>5 μm) were calculated from 10 randomly selected viewing fieldsTo identify SG specific modulators, SG cores were isolated according to the well-established procedures (Jain et al., 2016) from the TIA-1 overexpressing HeLa cells. These purified SG cores were divided into small aliquots and treated with compounds from a library containing 116 natural compounds isolated from 58 kinds of Chinese traditional medical plants (Fig. 1A). The library covers chemical compounds belonging to alkaloids, glycosides, ketones, flavonoids, phenylpropanoids, phenols, quinones, terpenoids, and steroids. After treatment, the effect of these compounds was determined by the dissociation of GFP-TIA-1 from SGs evaluated by the ratio of precipitation to supernatant fraction. Our results showed that 23 compounds facilitated SG dissociation, while 60 promoted SG assembly (Fig. 1B). Among them, 3 compounds could successfully decrease over 30% of the precipitation fraction, while 13 compounds increased the precipitation fraction above 2-fold. The library contains 13 benzene derivatives. A preliminary analysis suggested that two methoxy groups might be required for the SG dissociation ability (Fig. 1C). Further research using a larger library of benzene derivatives is needed to elucidate the structure-activity relationship. Among the effective compounds, most of them did not affect cell viability, while 4 of them showed significant cytotoxicity at a concentration of 10 μmol/L (Fig. 1D).We selected two highly effective non-cytotoxic compounds (Fig. 1B), C13 (syringic acid) and C85 (troxerutin), for further investigations. Western blot analysis indicated that C13 and C85 had opposite functions on SG stability (Fig. 1E). Consistently, confocal microscopy of the isolated SG cores treated with C13 showed a disperse GFP fluorescence pattern, which was caused by the re-dissolution of GFP-TIA-1 from SGs. Those treated with C85 had more larger SGs with size above 2 μm when compared with the control (Fig. 1F), implying that C85 could stabilize SGs and perturb the equilibrium between reversible SG assembly and disassembly. C13 and C85 were also very effective for SGs induced by TIA-1 overexpression or arsenite treatment in the HeLa cells (Fig. 2A). Quantitative analysis indicated that the percentage of cells with SGs induced by TIA-1 was significantly reduced by C13 and promoted by C85 in a concentration-dependent manner (Fig. 2B). As for SGs induced by arsenite treatment, C13 decreased both the number and relative mean fluorescence intensity of SGs, while C85 had the opposite effect (Fig. 2C). The addition of C13 or C85 did not affect the formation of protein aggregates formed by a disease-causing mutant αB-crystallin R120G visualized by the fused GFP or P bodies detected by a marker protein EDC4 (Fig. S1) or DCP1a (data not shown). This suggested that C13 and C85 were more likely to be specific modulators of SG formation but not the other types of cytoplasmic granules.Open image in new windowFigure 2Verification of the action of C13 and C85 on SGs in HeLa cells. (A) Representative confocal microscope images showing the effects of various concentrations of C13 and C85 on SGs in HeLa cells induced by overexpression of GFP-TIA-1 or 0.5 mmol/L arsenite for 30 min. Scare bar, 10 μm. Treatment of C13 or C85 did not affect the transient transfection efficiency or expression level of TIA-1 as evaluated by fluorescence mean intensity and Western-blot analysis (data not shown). (B) Quantitative analysis of the actions of C13 and C85 on GFP-TIA-1 overexpression-induced SGs in HeLa cells. The percentages of cells with SGs were calculated from 10 randomly selected viewing fields. (C) Quantitative analysis of the effects of C13 and C85 on the number of SGs per cell and mean fluorescence intensity. SGs were induced by 0.5 mmol/L arsenite for 30 min. **P < 0.01, ***P < 0.001. (D) Effect of the two compounds on cell survival when the HeLa cells were treated by various stressors. Cell viability was measured immediately after the stress treatment to reflect the sensitivity of stress response. (E) Cell death determined by flow cytometry analysis for samples double stained using annexin V and propidium iodide. (F) Effect of the two compounds on the recovery of the HeLa cells from stressed conditions. The cell viability was measured by refreshing the culture medium to remove the stressors and cultivated for 2 h. All cell viability data are presented as mean ± S.E.M. (n = 3)SGs can be induced by many stressors and the compositions may differ for SG induced by different stresses. Both compounds were also effective for SGs induced by H2O2, heat shock, DTT, TG and TM (Fig. S2A), suggesting that the compounds had a general effect on SG assembly/disassembly in the cells though they were screened using the isolated SG cores induced by TIA-1 overexpression. Quantitative analysis (Fig. S2B) indicated that C13 could decrease the percentage of cells containing SGs when the cells were treated with effective SG inducers such as arsenite, H2O2, and DTT. For mild SG-inducing conditions including heat shock, TG and TM treatments, C85 enhanced SG formation though C85 could not induce SG formation under normal conditions by itself (data not shown).Previous functional studies of SGs mainly performed using exogenous stressors or overexpressing/knocking down SG components. It is worth noting that these stressors/proteins may have pleiotropic cellular effects although it is clear that they are involved in SG formation (Mahboubi and Stochaj, 2017; Panas et al., 2016). Both C13 and C85 did not have cytotoxicity (Fig. 1D) and did not induce SG formation under non-stressed conditions (data not shown). Cell viability was further studied under conditions in the presence of stressors. Similar cell survival results were observed in HeLa and HEK 293A cells, and the data of HeLa cells are shown in Figs. 2D–F and S3. Time-course study was performed for cells transfected with GFP-C3 or GFP-TIA-1. The culture medium was not refreshed during cultivation and thereby cells will subject to starvation after 12 h cultivation. Compared to the control group, C13 greatly facilitated cell survival but C85 decreased cell viability after 8 h cultivation (Fig. S3A). Pre-treatment of the cells with the two compounds had dissimilar effects on cellular stress response to arsenite, H2O2, DTT, heat shock, TG and TM treatments (Fig. 2D and 2E). For all stressors, the SG-promoting compound C85 impaired cell survival and induce cell death. The SG-dissociating compound C13 slightly decreased cell viability for heat shock, TG and TM treatments, while enhanced cell survival and prevented cell death for arsenite, H2O2, and DTT treatments. The effects of both compounds showed a stressor dose dependency for arsenite and H2O2 treatments (Fig. S3B). It seems that SG inhibition by C13 affected cell survival differentially for various stresses, which might be caused by the dissimilar compositions of SGs induced by different stressors (Panas et al., 2016) and the severity of the stresses. When the stressors were removed by refreshing the culture medium of the cells, C85 showed ever greater impairments on cell viability, while C13 was beneficial to the cells recovered from all stressed conditions (Fig. 2F). The unappreciated effect of C85 on cell survival was stressor treating-time dependent, while C13 had similar protecting effect for the tested conditions (Fig. S3C).Formation of SGs has been proposed to facilitate stress response of the eukaryotic cells (Mahboubi and Stochaj, 2017; Protter and Parker, 2016). Although C85 was not cytotoxic under normal cultivating conditions, cells treated with C85 showed hypersensitive to various stressors. The SG-promoting compound C85 was deleterious to not only cell survival under stressed conditions but also recovery after stressors were removed. The extraordinarily enhanced SG formation by C85 might recruit essential stress-fighting mRNAs/proteins into SGs and thereby impair the cellular machines required for survival and recovery. The SG-dissociating compound C13 had complicated effect on cell stress response. Compared with the control group, cells treated with C13 were more resistant to arsenite, H2O2 and DTT, but was more sensitive to heat shock, TG, and TM. This implied that a proper equilibrium between SG assembly/disassembly was required for cellular stress response. Nonetheless, C13 facilitated the cells recovered from the stressed conditions, suggesting that a faster dissociation of SGs was beneficial to the survived cells to return to their normal states.In conclusion, herein we developed a simple and effective screening strategy to identify chemical compounds modulating SG assembly/disassembly. A number of natural aromatic compounds had been identified to have the potency to modulate SG formation. The action of two highly effective and nontoxic compounds, C13 (syringic acid) and C85 (troxerutin), were verified for various subtypes of SGs induced by dissimilar stressors. More importantly, both compounds did not affect the formation of other types of cytoplasmic protein aggregates or RNA granules, suggesting that these two compounds were highly SG-specific and could be used to modulate SG formation in the cells without modifications of the other types of cellular bodies. A preliminary functional study indicated that the SG-promoting compound C85 impaired both cellular stress response and recovery from stressed conditions. The SG-dissociating compound C13 had complicated effect on stress response but facilitate recovery after the cells were released from stressors. Our results suggested that there do have SG-specific modulators although the microscopic visible membraneless organelles share some common assembly/disassembly mechanism. Screening using a larger library combined with structural design will provide more SG-specific modulators with higher efficiency. Meanwhile, herein we only performed a preliminary functional study of SGs using these modulators. Further research is needed to elucidate the mechanism of these compounds and their cellular consequence by modulating SG assembly/disassembly.FOOTNOTESThis study was supported by funds from the China Postdoctoral Science Foundation (No. 158358) and State Key Laboratory of Membrane Biology (to Y.-B. Yan). Dr. X.-J. Chen was supported by the Excellent Postdoctoral fellowship from the Tsinghua-Peking Joint Center for Life Sciences.Li-Dan Hu, Xiang-Jun Chen, Xiao-Yan Liao and Yong-Bin Yan declare no conflict of interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.YBY conceived and designed the experiments. LDH, XJC, and XYL performed the experiments. LDH, XJC, and YBY analyzed the data. LDH and YBY wrote the paper.
Co-reporter:Liang-Bo Qi;Li-Dan Hu;Huihui Liu;Hai-Yun Li;Xiao-Yao Leng
Protein & Cell 2016 Volume 7( Issue 7) pp:501-515
Publication Date(Web):2016 July
DOI:10.1007/s13238-016-0284-3
β/γ-Crystallins are predominant structural proteins in the cytoplasm of lens fiber cells and share a similar fold composing of four Greek-key motifs divided into two domains. Numerous cataract-causing mutations have been identified in various β/γ-crystallins, but the mechanisms underlying cataract caused by most mutations remains uncharacterized. The S228P mutation in βB1-crystallin has been linked to autosomal dominant congenital nuclear cataract. Here we found that the S228P mutant was prone to aggregate and degrade in both of the human and E. coli cells. The intracellular S228P aggregates could be redissolved by lanosterol. The S228P mutation modified the refolding pathway of βB1-crystallin by affecting the formation of the dimeric intermediate but not the monomeric intermediate. Compared with native βB1-crystallin, the refolded S228P protein had less packed structures, unquenched Trp fluorophores and increased hydrophobic exposure. The refolded S228P protein was prone to aggregate at the physiological temperature and decreased the protective effect of βB1-crystallin on βA3-crystallin. Molecular dynamic simulation studies indicated that the mutation decreased the subunit binding energy and modified the distribution of surface electrostatic potentials. More importantly, the mutation separated two interacting loops in the C-terminal domain, which shielded the hydrophobic core from solvent in native βB1-crystallin. These two interacting loops are highly conserved in both of the N- and C-terminal domains of all β/γ-crystallins. We propose that these two interacting loops play an important role in the folding and structural stability of β/γ-crystallin domains by protecting the hydrophobic core from solvent access.
Co-reporter:Xiao-Yao Leng, Sha Wang, Ni-Qian Cao, Liang-Bo Qi, and Yong-Bin Yan
Biochemistry 2014 Volume 53(Issue 15) pp:
Publication Date(Web):March 26, 2014
DOI:10.1021/bi500146d
β/γ-Crystallins are the major structural proteins in mammalian lens. The N-terminal truncation of βB1-crystallin has been associated with the regulation of β-crystallin size distributions in human lens. Herein we studied the roles of βB1 N-terminal extension in protein structure and folding by constructing five N-terminal truncated forms. The truncations did not affect the secondary and tertiary structures of the main body as well as stability against denaturation. Truncations with more than 28 residues off the N-terminus promoted the dissociation of the dimeric βB1 into monomers in diluted solutions. Interestingly, the N-terminal extension facilitated βB1 to adopt the correct folding pathway, while truncated proteins were prone to undergo the misfolding/aggregation pathway during kinetic refolding. The N-terminal extension of βB1 acted as an intramolecular chaperone (IMC) to regulate the kinetic partitioning between folding and misfolding. The IMC function of the N-terminal extension was also critical to the correct refolding of β-crystallin heteromer and the action of the lens-specific molecular chaperone αA-crystallin. The cooperation between IMC and molecular chaperones produced a much stronger chaperoning effect than if they acted separately. To our knowledge, this is the first report showing the cooperation between IMC and molecular chaperones.
Co-reporter:Sha Wang, Xiao-Yao Leng, and Yong-Bin Yan
Biochemistry 2011 Volume 50(Issue 48) pp:
Publication Date(Web):October 27, 2011
DOI:10.1021/bi201375p
β-Crystallins are the major structural proteins in mammalian lens, and their stability is critical in maintaining the transparency and refraction index of the lens. Among the seven β-crystallins, βA3-crystallin and βB1-crystallin, an acidic and a basic β-crystallin, respectively, can form heteromers in vivo. However, the physiological roles of the heteromer have not been fully elucidated. In this research, we studied whether the basic β-crystallin facilitates the folding of acidic β-crystallin. Equilibrium folding studies revealed that the βA3-crystallin and βB1-crystallin homomers and the βA3/βB1-crystallin heteromer all undergo similar five-state folding pathways which include one dimeric and two monomeric intermediates. βA3-Crystallin was found to be the most unstable among the three proteins, and the transition curve of βA3/βB1-crystallin was close to that of βB1-crystallin. The dimeric intermediate may be a critical determinant in the aggregation process and thus is crucial to the lifelong stability of the β-crystallins. A comparison of the Gibbs free energy of the equilibrium folding suggested that the formation of heteromer contributed to the stabilization of the dimer interface. On the other hand, βA3-crystallin, the only protein whose refolding is challenged by serious aggregation, can be protected by βB1-crystallin in a dose-dependent manner during the kinetic co-refolding. However, the protection is not observed in the presence of the pre-existed well-folded βB1-crystallin. These findings suggested that the formation of β-crystallin heteromers not only stabilizes the unstable acidic β-crystallin but also protects them against aggregation during refolding from the stress-denatured states.
Co-reporter:Min Pang, Jing-Tan Su, Shan Feng, Zhi-Wei Tang, Feng Gu, Meng Zhang, Xu Ma, Yong-Bin Yan
Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 2010 Volume 1804(Issue 4) pp:948-956
Publication Date(Web):April 2010
DOI:10.1016/j.bbapap.2010.01.001
α-crystallin is a molecular chaperone that maintains the optical properties of the lens and delays the onset scattering caused by aging-related protein aggregation. In this research, we found that the missense mutation R116H resulted in an altered size distribution, impaired packing of the secondary structures and modified quaternary structure with great hydrophobic exposure. The mutant exhibited a substrate-dependent chaperone (aggregation–inhibition) or anti-chaperone (aggregation–promotion) effect. Equilibrium unfolding experiments indicated that the mutation stabilized an aggregation-prone intermediate which was not populated during the unfolding of the wild-type protein. The accumulation of this intermediate greatly promoted the formation of non-native large oligomers or aggregates during unfolding. These results suggested that both the aggregation of the mutant upon stress and co-deposition with the target proteins were likely to be responsible for the onset of cataract.
Co-reporter:Shan Feng, Zhen Xu, Yong-Bin Yan
Journal of Inorganic Biochemistry 2008 Volume 102(Issue 4) pp:928-935
Publication Date(Web):April 2008
DOI:10.1016/j.jinorgbio.2007.12.013
Creatine kinase (CK) is a key enzyme to maintain the energy homeostasis in vertebrate excitable tissues. Due to its importance in cellular energetics, the activity and level of CK are crucial to cellular and body functions. CK is sensitive to oxidative stresses and is thought to be one of the main targets of oxidative modification in neurodegenerative diseases. In this research, we investigated the effect of copper, an essential trace element for all organisms and an inducer of the reactive oxygen species, on CK refolding. It was found that trace amounts of Cu2+ (3 mol eq of Cu2+) could efficiently block the refolding of CK. The Cu2+-trapped CK could not be reactivated by the addition of EDTA, but could be reactivated by DTT. Spectroscopic experiments suggested that copper ions blocked CK refolding by specifically binding with the monomeric refolding intermediate, which further retarded CK refolding and promoted the formation of off-pathway aggregates. The results herein suggested that Cu2+-induced CK dysfunction might be caused not only by the post-translational oxidation, but also by the direct binding of copper ions with the newly-synthesized polypeptides.
Co-reporter:Sha Wang, Wei-Feng Liu, Yong-Zhi He, Ao Zhang, Li Huang, Zhi-Yang Dong, Yong-Bin Yan
Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 2008 Volume 1784(Issue 3) pp:445-454
Publication Date(Web):March 2008
DOI:10.1016/j.bbapap.2007.12.001
Superoxide dismutases (SODs), which are the first line of cellular defense against the toxic effects of reactive oxygen species, are metalloenzymes that catalyze the disproportionation of superoxide radicals to produce oxygen and hydrogen peroxide. Although much effort has been devoted to the folding mechanisms of Cu/Zn-SODs, little is known about the folding of Fe-SODs. In this research, the equilibrium unfolding and refolding of TcSOD, a tetrameric hyperthermostable Fe-SOD, were investigated by circular dichroism, intrinsic fluorescence, ANS fluorescence, size-exclusion chromatography and cross-linking experiments. The results herein suggested that the guanidine hydrochloride-induced unfolding of TcSOD involved a stable monomeric intermediate and a possible tetrameric intermediate. The Gibbs free energy of TcSOD dissociation was about 3-fold larger than that of the monomeric intermediate unfolding, which suggested that the quaternary structure plays a crucial role in TcSOD stability. A comparison of the thermodynamic parameters between TcSOD and other SODs also suggested that the stability of quaternary structure might be responsible for the hyperthemostability of TcSOD.
Co-reporter:Wei-Feng Liu, Yong-Bin Yan
Protein Expression and Purification (July 2008) Volume 60(Issue 1) pp:46-52
Publication Date(Web):1 July 2008
DOI:10.1016/j.pep.2008.03.008
Pop2, a component of the Ccr4–Not complex, functions as a deadenylase both in vitro and in vivo. In this research, we found that the recombinant human Pop2 (hPop2) mainly existed in a compact monomeric state with a α + β tertiary structure type. The percentages of the secondary structures evaluated from the CD spectrum were about 37% α-helix, 14% β-sheet, and 19% β-turns. The optimal condition for hPop2 catalysis was pH 7–8 at 37 °C. Mg2+, Mn2+, and Co2+ had similar effects on the deadenylation activity of hPop2, and the optimal concentration was 0.3–0.5 mM. The deadenylase activity of hPop2 was, at least partially, specific when coordinated with divalent metal ions. The enzyme was not inhibited much by the nucleotide analogs, and the product 5′-AMP was the most efficient inhibitor. The dissimilarity in the metal ion dependence and inhibitory effects of the nucleotide analogs suggested that various deadenylases might have differential regulation mechanisms.
Co-reporter:Xiang-Jun Chen, Xin-Hang Zhang, Li-Dan Hu, Jia-Quan Zhang, Yan Jiang, Yi Yang, Yong-Bin Yan
International Journal of Biological Macromolecules (January 2016) Volume 82() pp:369-374
Publication Date(Web):1 January 2016
DOI:10.1016/j.ijbiomac.2015.10.015
•The deadenylase gene Dscaf1 is cloned from the halotolerant alga Dunaliella salina.•Dscaf1 promoter contains many cis-acting elements responsive to abiotic stress.•Dscaf1 is highly expressed under hypersaline conditions.•Dscaf1 exhibits two types of two-stage responses to environmental stresses.Regulation of mRNA fates can alter the gene-expression profile promptly and specifically. Deadenylation is a process of which the poly(A) tail is degraded by deadenylases. By regulating mRNA turnover, deadenylases are involved in various vital cellular processes including stress responses. However, it is unclear whether deadenylases play a role in the adaption/tolerance of extremophiles. In this research, we cloned the deadenylase caf1 from Dunaliella salina (dscaf1), a unicellular green alga with exceptional halotolerance. In silicon analysis indicated that compared with the mesophilic alga Chlamydomonas reinhardtii caf1, dscaf1 promoter contained more elements responsive to abiotic stresses. Dscaf1 had an extremely high expression level under hypersaline conditions. When the D. salina cells were subject to stress shock, a two-stage response was observed for dscaf1 expression. The mRNA level of dscaf1 had an immediate 2–4 fold increase and followed by an ∼10 fold increase after hyperosmotic, heat or UV treatment, while had an about 3 fold increase quickly followed by an abrupt decrease after hypoosmotic or cold shock. The dissimilarity in dscaf1 expression patterns suggested that DsCaf1 is a stress-responsive deadenylase with the ability to regulate fates of a specific group of mRNAs for a certain type of stress.
Co-reporter:Li-Na Zhang, Yong-Bin Yan
Biochimica et Biophysica Acta (BBA) - Molecular Cell Research (February 2015) Volume 1853(Issue 2) pp:522-534
Publication Date(Web):February 2015
DOI:10.1016/j.bbamcr.2014.12.004
Co-reporter:Shan Feng, Zhen Xu, Yong-Bin Yan
Journal of Inorganic Biochemistry (April 2008) Volume 102(Issue 4) pp:928-935
Publication Date(Web):1 April 2008
DOI:10.1016/j.jinorgbio.2007.12.013
Creatine kinase (CK) is a key enzyme to maintain the energy homeostasis in vertebrate excitable tissues. Due to its importance in cellular energetics, the activity and level of CK are crucial to cellular and body functions. CK is sensitive to oxidative stresses and is thought to be one of the main targets of oxidative modification in neurodegenerative diseases. In this research, we investigated the effect of copper, an essential trace element for all organisms and an inducer of the reactive oxygen species, on CK refolding. It was found that trace amounts of Cu2+ (3 mol eq of Cu2+) could efficiently block the refolding of CK. The Cu2+-trapped CK could not be reactivated by the addition of EDTA, but could be reactivated by DTT. Spectroscopic experiments suggested that copper ions blocked CK refolding by specifically binding with the monomeric refolding intermediate, which further retarded CK refolding and promoted the formation of off-pathway aggregates. The results herein suggested that Cu2+-induced CK dysfunction might be caused not only by the post-translational oxidation, but also by the direct binding of copper ions with the newly-synthesized polypeptides.
Co-reporter:Zhe Chen, Xiang-Jun Chen, Mengdie Xia, Hua-Wei He, Sha Wang, Huihui Liu, Haipeng Gong, Yong-Bin Yan
Biophysical Journal (8 August 2012) Volume 103(Issue 3) pp:
Publication Date(Web):8 August 2012
DOI:10.1016/j.bpj.2012.07.002
Intramolecular chaperones (IMCs), which are specific domains/segments encoded in the primary structure of proteins, exhibit chaperone-like activity against the aggregation of the other domains in the same molecule. In this research, we found that the truncation of the linker greatly promoted the thermal aggregation of the isolated C-terminal domain (CTD) of rabbit muscle creatine kinase (RMCK). Either the existence of the linker covalently linked to CTD or the supply of the synthetic linker peptide additionally could successfully protect the CTD of RMCK against aggregation in a concentration-dependent manner. Truncated fragments of the linker also behaved as a chaperone-like effect with lower efficiency, revealing the importance of its C-terminal half in the IMC function of the linker. The aggregation sites in the CTD of RMCK were identified by molecular dynamics simulations. Mutational analysis of the three key hydrophobic residues resulted in opposing effects on the thermal aggregation between the CTD with intact or partial linker, confirming the role of linker as a lid to protect the hydrophobic residues against exposure to solvent. These observations suggested that the linkers in multidomain proteins could act as IMCs to facilitate the correct folding of the aggregation-prone domains. Furthermore, the intactness of the IMC linker after proteolysis modulates the production of off-pathway aggregates, which may be important to the onset of some diseases caused by the toxic effects of aggregated proteolytic fragments.
Co-reporter:Wei-Feng Liu, Ao Zhang, Yuan Cheng, Hai-Meng Zhou, Yong-Bin Yan
FEBS Letters (6 March 2007) Volume 581(Issue 5) pp:1047-1052
Publication Date(Web):6 March 2007
DOI:10.1016/j.febslet.2007.02.008
Poly(A)-specific ribonuclease (PARN), a member of the DEDD family, is a key enzyme involved in the deadenylation of mRNA in higher eukaryotic cells. In this research, it was found that Mg2+ could protect PARN against thermal inactivation by increasing the midpoint of inactivation and decreasing the inactivation rate. This protective effect was unique to Mg2+ in a concentration-dependent manner. However, the thermal unfolding and aggregation was promoted by the addition of Mg2+ at high temperatures. These results revealed that Mg2+ might have dual effects on PARN stability: protecting the active site but endangering the overall structural stability.
Co-reporter:Ao Zhang, Wei-Feng Liu, Yong-Bin Yan
Archives of Biochemistry and Biophysics (15 May 2007) Volume 461(Issue 2) pp:255-262
Publication Date(Web):15 May 2007
DOI:10.1016/j.abb.2007.02.023
Co-reporter:Jing-Tan Su, Sung-Hye Kim, Yong-Bin Yan
Biophysical Journal (15 January 2007) Volume 92(Issue 2) pp:
Publication Date(Web):15 January 2007
DOI:10.1529/biophysj.106.093666
Aminoacylase I (ACYI) catalyzes the stereospecific hydrolysis of L-acylamino acids and is generally assumed to be involved in the final step of the degradation of intracellular N-acetylated proteins. Apart from its crucial functions in intracellular amino acid metabolism, ACYI also has substantial commercial importance for the optical resolution of N-acylated DL-amino acids. As a zinc-dependent enzyme, ACYI is quite stable against heat-induced denaturation and can be regarded as a thermostable enzyme with an optimal temperature for activity of ∼65°C. In this research, the sequential events in ACYI thermal denaturation were investigated by a combination of spectroscopic methods and related resolution-enhancing techniques. Interestingly, the results from fluorescence and infrared (IR) spectroscopy clearly indicated that a pretransitional stage existed at temperatures from 50°C to 66°C. The thermal unfolding of ACYI might be a three-state process involving an aggregation-prone intermediate appearing at ∼68°C. The pretransitional structural changes involved the partial unfolding of the solvent-exposed β-sheet structures and the transformation of about half of the Class I Trp fluorophores to Class II. Our results also suggested that the usage of resolution-enhancing techniques could provide valuable information of the step-wise unfolding of proteins.

![3,5,9-Trioxa-4-phosphapentacosan-1-aminium,4-hydroxy-N,N,N-trimethyl-10-oxo-7-[(1-oxohexadecyl)oxy]-, inner salt, 4-oxide](http://img.cochemist.com/ccimg/2700/2644-64-6.png)
![3,5,9-Trioxa-4-phosphapentacosan-1-aminium,4-hydroxy-N,N,N-trimethyl-10-oxo-7-[(1-oxohexadecyl)oxy]-, inner salt, 4-oxide](http://img.cochemist.com/ccimg/2700/2644-64-6_b.png)












![(4S)-4,9-DIHYDROXY-2,2-DIMETHYL-3,4-DIHYDRO-2H-BENZO[G]CHROMENE-5<WBR />,10-DIONE](http://img.cochemist.com/ccimg/3100/3036-82-6.png)
![(4S)-4,9-DIHYDROXY-2,2-DIMETHYL-3,4-DIHYDRO-2H-BENZO[G]CHROMENE-5<WBR />,10-DIONE](http://img.cochemist.com/ccimg/3100/3036-82-6_b.png)