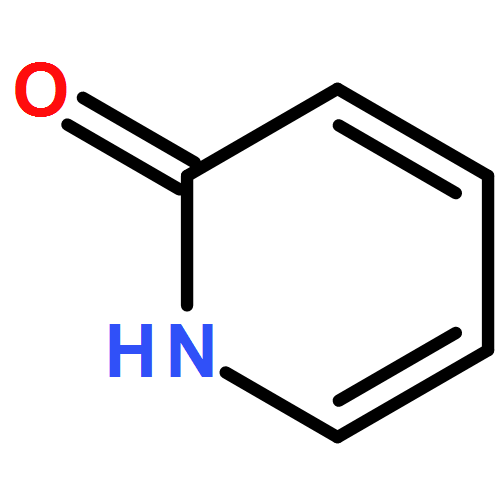
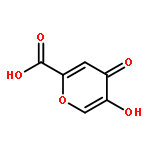
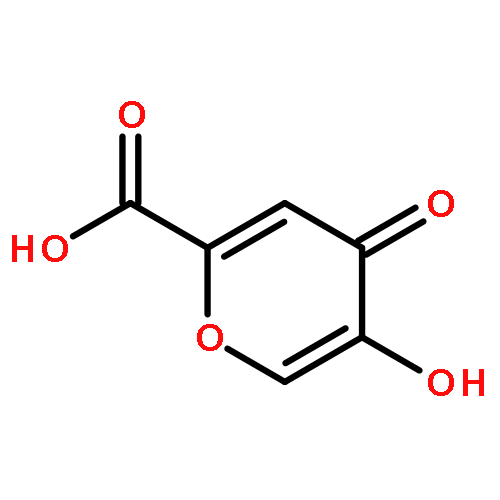
Co-reporter:Meaghan E. Germain and Michael J. Knapp
Inorganic Chemistry November 3, 2008 Volume 47(Issue 21) pp:9748-9750
Publication Date(Web):October 2, 2008
DOI:10.1021/ic801317x
Peroxide-based explosives, like triacetone triperoxide (TATP), are important targets for detection because of their broad use in improvised explosives but pose challenges. We report a highly sensitive turn-on fluorescence detection for H2O2 and organic peroxides, including TATP. The detection strategy relies on oxidative deboronation to unmask H2Salen, which subsequently binds Zn2+ to form fluorescent Zn(Salen). Sensitivity is excellent, with detection limits below 10 nM for H2O2, TATP, and benzoyl peroxide. In addition, acid treatment is necessary to sense TATP, suggesting the potential to discriminate between H2O2 and TATP based upon minimal sample pretreatment.
Co-reporter:John A. Hangasky, Cornelius Y. Taabazuing, Cristina B. Martin, Scott J. Eron, Michael J. Knapp
Journal of Inorganic Biochemistry 2017 Volume 166() pp:26-33
Publication Date(Web):January 2017
DOI:10.1016/j.jinorgbio.2016.10.007
•The enzyme factor-inhibiting hypoxia inducible factor-1α is known as FIH.•A His2-Asp facial triad is a common motif in α-ketoglutarate-dependent oxygenases.•The Asp structurally links O2 and substrate binding sites.•Replacing the Asp ligand leads to uncoupled O2 activation.•The Asp ligand is essential for tight coupling in FIH.The factor inhibiting hypoxia inducible factor-1α (FIH) is a nonheme Fe(II)/αKG oxygenase using a 2-His-1-Asp facial triad. FIH activates O2 via oxidative decarboxylation of α-ketoglutarate (αKG) to generate an enzyme-based oxidant which hydroxylates the Asn803 residue within the C-terminal transactivation domain (CTAD) of HIF-1α. Tight coupling of these two sequential reactions requires a structural linkage between the Fe(II) and the substrate binding site to ensure that O2 activation occurs after substrate binds. We tested the hypothesis that the facial triad carboxylate (Asp201) of FIH linked substrate binding and O2 binding sites. Asp201 variants of FIH were constructed and thoroughly characterized in vitro using steady-state kinetics, crystallography, autohydroxylation, and coupling measurements. Our studies revealed each variant activated O2 with a catalytic efficiency similar to that of wild-type (WT) FIH (kcataKM(O2) = 0.17 μM− 1min− 1), but led to defects in the coupling of O2 activation to substrate hydroxylation. Steady-state kinetics showed similar catalytic efficiencies for hydroxylation by WT-FIH (kcat/KM(CTAD) = 0.42 μM− 1 min− 1) and D201G (kcat/KM(CTAD) = 0.34 μM− 1 min− 1); hydroxylation by D201E was greatly impaired, while hydroxylation by D201A was undetectable. Analysis of the crystal structure of the D201E variant revealed steric crowding near the diffusible ligand site supporting a role for sterics from the facial triad carboxylate in the O2 binding order. Our data support a model in which the facial triad carboxylate Asp201 provides both steric and polar contacts to favor O2 access to the Fe(II) only after substrate binds, leading to coupled turnover in FIH and other αKG oxygenases.The facial triad carboxylate structurally links O2 and substrate binding in the factor-inhibiting hypoxia inducible factor-1α (FIH), an α-ketoglutarate dependent oxygenase.
Co-reporter:Cornelius Y. Taabazuing, Justin Fermann, Scott Garman, and Michael J. Knapp
Biochemistry 2016 Volume 55(Issue 2) pp:277-286
Publication Date(Web):January 5, 2016
DOI:10.1021/acs.biochem.5b01003
The Fe2+/α-ketoglutarate (αKG)-dependent oxygenases use molecular oxygen to conduct a wide variety of reactions with important biological implications, such as DNA base excision repair, histone demethylation, and the cellular hypoxia response. These enzymes follow a sequential mechanism in which O2 binds and reacts after the primary substrate binds, making those structural factors that promote productive O2 binding central to their chemistry. A large challenge in this field is to identify strategies that engender productive turnover. Factor inhibiting HIF (FIH) is a Fe2+/αKG-dependent oxygenase that forms part of the O2 sensing machinery in human cells by hydroxylating the C-terminal transactivation domain (CTAD) found within the HIF-1α protein. The structure of FIH was determined with the O2 analogue NO bound to Fe, offering the first direct insight into the gas binding geometry in this enzyme. Through a combination of density functional theory calculations, {FeNO}7 electron paramagnetic resonance spectroscopy, and ultraviolet–visible absorption spectroscopy, we demonstrate that CTAD binding stimulates O2 reactivity by altering the orientation of the bound gas molecule. Although unliganded FIH binds NO with moderate affinity, the bound gas can adopt either of two orientations with similar stability; upon CTAD binding, NO adopts a single preferred orientation that is appropriate for supporting oxidative decarboxylation. Combined with other studies of related enzymes, our data suggest that substrate-induced reorientation of bound O2 is the mechanism utilized by the αKG oxygenases to tightly couple O2 activation to substrate hydroxylation.
Co-reporter:Serap Pektas, Cornelius Y. Taabazuing, and Michael J. Knapp
Biochemistry 2015 Volume 54(Issue 18) pp:2851-2857
Publication Date(Web):April 10, 2015
DOI:10.1021/bi501540c
PHD2 is a 2-oxoglutarate, non-heme Fe2+-dependent oxygenase that senses O2 levels in human cells by hydroxylating two prolyl residues in the oxygen-dependent degradation domain (ODD) of HIF1α. Identifying the active site contacts that determine the rate of reaction at limiting O2 concentrations is crucial for understanding how this enzyme senses pO2 and may suggest methods for chemically altering hypoxia responses. A hydrogen bonding network extends from the Fe(II) cofactor through ordered waters to the Thr387 residue in the second coordination sphere. Here we tested the impact of the side chain of Thr387 on the reactivity of PHD2 toward O2 through a combination of point mutagenesis, steady state kinetic experiments and {FeNO}7 EPR spectroscopy. The steady state kinetic parameters for Thr387 → Asn were very similar to those of wild-type (WT) PHD2, but kcat and kcat/KM(O2) for Thr387 → Ala were increased by roughly 15-fold. X-Band electron paramagnetic resonance spectroscopy of the {FeNO}7 centers of the (Fe+NO+2OG) enzyme forms showed the presence of a more rhombic line shape in Thr387 → Ala than in WT PHD2, indicating an altered conformation for bound gas in this variant. Here we show that the side chain of residue Thr387 plays a significant role in determining the rate of turnover by PHD2 at low O2 concentrations.
Co-reporter:Cornelius Y. Taabazuing, John A. Hangasky, Michael J. Knapp
Journal of Inorganic Biochemistry 2014 Volume 133() pp:63-72
Publication Date(Web):April 2014
DOI:10.1016/j.jinorgbio.2013.12.010
The ability to sense and adapt to changes in pO2 is crucial for basic metabolism in most organisms, leading to elaborate pathways for sensing hypoxia (low pO2). This review focuses on the mechanisms utilized by mammals and bacteria to sense hypoxia. While responses to acute hypoxia in mammalian tissues lead to altered vascular tension, the molecular mechanism of signal transduction is not well understood. In contrast, chronic hypoxia evokes cellular responses that lead to transcriptional changes mediated by the hypoxia inducible factor (HIF), which is directly controlled by post-translational hydroxylation of HIF by the non-heme Fe(II)/αKG-dependent enzymes FIH and PHD2. Research on PHD2 and FIH is focused on developing inhibitors and understanding the links between HIF binding and the O2 reaction in these enzymes. Sulfur speciation is a putative mechanism for acute O2-sensing, with special focus on the role of H2S. This sulfur-centered model is discussed, as are some of the directions for further refinement of this model. In contrast to mammals, bacterial O2-sensing relies on protein cofactors that either bind O2 or oxidatively decompose. The sensing modality for bacterial O2-sensors is either via altered DNA binding affinity of the sensory protein, or else due to the actions of a two-component signaling cascade. Emerging data suggests that proteins containing a hemerythrin-domain, such as FBXL5, may serve to connect iron sensing to O2-sensing in both bacteria and humans. As specific molecular machinery becomes identified, these hypoxia sensing pathways present therapeutic targets for diseases including ischemia, cancer, or bacterial infection.Hypoxia sensing follows multiple avenues, ranging from cellular responses to changes in vascular tension. A number of molecular players are implicated, including gasogens, sulfur metabolites, and ions. Defining these molecular interactions is key to discovering new biochemistry, and holds promise for future therapeutic targets.
Co-reporter:John A. Hangasky, Geoffrey T. Ivison, and Michael J. Knapp
Biochemistry 2014 Volume 53(Issue 36) pp:
Publication Date(Web):August 13, 2014
DOI:10.1021/bi500703s
Nonheme Fe(II)/αKG-dependent oxygenases catalyze diverse reactions, typically inserting an O atom from O2 into a C–H bond. Although the key to their catalytic cycle is the fact that binding and positioning of primary substrate precede O2 activation, the means by which substrate binding stimulates turnover is not well understood. Factor Inhibiting HIF (FIH) is a Fe(II)/αKG-dependent oxygenase that acts as a cellular oxygen sensor in humans by hydroxylating the target residue Asn803, found in the C-terminal transactivation domain (CTAD) of hypoxia inducible factor-1. FIH-Gln239 makes two hydrogen bonds with CTAD-Asn803, positioning this target residue over the Fe(II). We hypothesized the positioning of the side chain of CTAD-Asn803 by FIH-Gln239 was critical for stimulating O2 activation and subsequent substrate hydroxylation. The steady-state characterization of five FIH-Gln239 variants (Ala, Asn, Glu, His, and Leu) tested the role of hydrogen bonding potential and sterics near the target residue. Each variant exhibited a 20–1200-fold decrease in kcat and kcat/KM(CTAD), but no change in KM(CTAD), indicating that the step after CTAD binding was affected by point mutation. Uncoupled O2 activation was prominent in these variants, as shown by large coupling ratios (C = [succinate]/[CTAD-OH] = 3–5) for each of the FIH-Gln239 → X variants. The coupling ratios decreased in D2O, indicating an isotope-sensitive inactivation for variants, not observed in the wild type. The data presented indicate that the proper positioning of CTAD-Asn803 by FIH-Gln239 is necessary to suppress uncoupled turnover and to support substrate hydroxylation, suggesting substrate positioning may be crucial for directing O2 reactivity within the broader class of αKG hydroxylases.
Co-reporter:John A. Hangasky, Hasand Gandhi, Meaghan A. Valliere, Nathaniel E. Ostrom, and Michael J. Knapp
Biochemistry 2014 Volume 53(Issue 51) pp:
Publication Date(Web):November 25, 2014
DOI:10.1021/bi501246v
Factor inhibiting HIF (FIH) is a cellular O2-sensing enzyme, which hydroxylates the hypoxia inducible factor-1α. Previously reported inverse solvent kinetic isotope effects indicated that FIH limits its overall turnover through an O2 activation step (Hangasky, J. A., Saban, E., and Knapp, M. J. (2013) Biochemistry52, 1594−1602). Here we characterize the rate-limiting step for O2 activation by FIH using a suite of mechanistic probes on the second order rate constant kcat/KM(O2). Steady-state kinetics showed that the rate constant for O2 activation was slow (kcat/KM(O2)app = 3500 M–1 s–1) compared with other non-heme iron oxygenases, and solvent viscosity assays further excluded diffusional encounter with O2 from being rate limiting on kcat/KM(O2). Competitive oxygen-18 kinetic isotope effect measurements (18kcat/KM(O2) = 1.0114(5)) indicated that the transition state for O2 activation resembled a cyclic peroxohemiketal, which precedes the formation of the ferryl intermediate observed in related enzymes. We interpret this data to indicate that FIH limits its overall activity at the point of the nucleophilic attack of Fe-bound O2— on the C-2 carbon of αKG. Overall, these results show that FIH follows the consensus mechanism for αKG oxygenases, suggesting that FIH may be an ideal enzyme to directly access steps involved in O2 activation among the broad family of αKG oxygenases.
Co-reporter:John A. Hangasky, Cornelius Y. Taabazuing, Meaghan A. Valliere and Michael J. Knapp
Metallomics 2013 vol. 5(Issue 4) pp:287-301
Publication Date(Web):24 Jan 2013
DOI:10.1039/C3MT20153H
The Fe(II)/αketoglutarate (αKG) dependent oxygenases catalyze a diverse range of reactions significant in biological processes such as antibiotic biosynthesis, lipid metabolism, oxygen sensing, and DNA and RNA repair. Although functionally diverse, the eight-stranded β-barrel (cupin) and HX(D/E)XnH facial triad motifs are conserved in this super-family of enzymes. Crystal structure analysis of 25 αKG oxygenases reveals two stereoisomers of the Fe cofactor, Anti and Clock, which differ in the relative position of the exchangeable ligand position and the primary substrate. Herein, we discuss the relationship between the chemical mechanism and the secondary coordination sphere of the αKG oxygenases, within the constraints of the stereochemistry of the Fe cofactor. Sequence analysis of the cupin barrel indicates that a small subset of positions constitute the second coordination sphere, which has significant ramifications for the structure of the ferryl intermediate. The competence of both Anti and Clock stereoisomers of Fe points to a ferryl intermediate that is 5 coordinate. The small number of conserved close contacts within the active sites of αKG oxygenases can be extended to chemically related enzymes, such as the αKG-dependent halogenases SyrB2 and CytC3, and the non-αKG dependent dioxygenases isopenicillin N synthase (IPNS) and cysteine dioxygenase (CDO).
Co-reporter:Serap Pektas, Michael J. Knapp
Journal of Inorganic Biochemistry 2013 Volume 126() pp:55-60
Publication Date(Web):September 2013
DOI:10.1016/j.jinorgbio.2013.05.006
•PHD2 prefers the CODD substrate by 20-fold over the NODD substrate.•Substrate selectivity can be explained by the relative rate constants.•Electrostatics accounts for a small part of the substrate selectivity.•Both NODD and CODD induce the same conformational change in PHD2.HIF prolyl-4-hydroxylase 2 (PHD2) is a non-heme Fe, 2-oxoglutarate (2OG) dependent dioxygenase that regulates the hypoxia inducible transcription factor (HIF) by hydroxylating two conserved prolyl residues in N-terminal oxygen degradation domain (NODD) and C-terminal oxygen degradation domain (CODD) of HIF-1α. Prior studies have suggested that the substrate preference of PHD2 arises from binding contacts with the β2β3 loop of PHD2. In this study we tested the substrate selectivity of PHD2 by kinetic competition assays, varied ionic strength, and global protein flexibility using amide H/D exchange (HDX). Our results revealed that PHD2 preferred CODD by 20-fold over NODD and that electrostatics influenced this effect. Global HDX monitored by mass spectrometry indicated that binding of Fe(II) and 2OG stabilized the overall protein structure but the saturating concentrations of either NODD or CODD caused an identical change in protein flexibility. These observations imply that both substrates stabilize the β2β3 loop to the same extent. Under unsaturated substrate conditions NODD led to a higher HDX rate than CODD due to its lower binding affinity to PHD2. Our results suggest that loop closure is the dominant contributor to substrate selectivity in PHD2.Electrostatics favors binding and loop closure for PHD2.
Co-reporter:John A. Hangasky, Evren Saban, and Michael J. Knapp
Biochemistry 2013 Volume 52(Issue 9) pp:
Publication Date(Web):January 25, 2013
DOI:10.1021/bi3015482
Oxygen homeostasis plays a critical role in angiogenesis, erythropoiesis, and cell metabolism. Oxygen homeostasis is set by the hypoxia inducible factor-1α (HIF-1α) pathway, which is controlled by factor inhibiting HIF-1α (FIH). FIH is a non-heme Fe(II), α-ketoglutarate (αKG)-dependent dioxygenase that inhibits HIF-1α by hydroxylating the C-terminal transactivation domain (CTAD) of HIF-1α at HIF-Asn803. A tight coupling between CTAD binding and O2 activation is essential for hypoxia sensing, making changes in the coordination geometry of Fe(II) upon CTAD encounter a crucial feature of this enzyme. Although the consensus chemical mechanism for FIH proposes that CTAD binding triggers O2 activation by causing the Fe(II) cofactor to release an aquo ligand, experimental evidence of this has been absent. More broadly, this proposed coordination change at Fe(II) has not been observed during steady-state turnover in any αKG oxygenase to date. In this work, solvent isotope effects (SIEs) were used as a direct mechanistic probe of substrate-triggered aquo release in FIH, as inverse SIEs (SIE < 1) are signatures for pre-equilibrium aquo release from metal ions. Our mechanistic studies of FIH have revealed inverse solvent isotope effects in the steady-state rate constants at limiting concentrations of CTAD or αKG [D2Okcat/KM(CTAD) = 0.40 ± 0.07, and D2Okcat/KM(αKG) = 0.32 ± 0.08], providing direct evidence of aquo release during steady-state turnover. Furthermore, the SIE at saturating concentrations of CTAD and αKG was inverse (D2Okcat = 0.51 ± 0.07), indicating that aquo release occurs after CTAD binds. The inverse kinetic SIEs observed in the steady state for FIH can be explained by a strong Fe–OH2 bond. The stable Fe–OH2 bond plays an important part in FIH’s regulatory role over O2 homeostasis in humans and points toward a strategy for tightly coupling O2 activation with CTAD hydroxylation that relies on substrate triggering.
Co-reporter:Shannon C. Flagg, Nitai Giri, Serap Pektas, Michael J. Maroney, and Michael J. Knapp
Biochemistry 2012 Volume 51(Issue 33) pp:
Publication Date(Web):June 29, 2012
DOI:10.1021/bi300229y
Prolyl hydroxylase domain 2 (PHD2) is deemed a primary oxygen sensor in humans, yet many details of its underlying mechanism are still not fully understood. (Fe2+ + αKG)PHD2 is 6-coordinate, with a 2His/1Asp facial triad occupying three coordination sites, a bidentate α-ketoglutarate occupying two sites, and an aquo ligand in the final site. Turnover is thought to be initiated upon release of the aquo ligand, creating a site for O2 to bind at the iron. Herein we show that steady-state turnover is faster under acidic conditions, with kcat exhibiting a kinetic pKa = 7.22. A variety of spectroscopic probes were employed to identify the active-site acid, through comparison of (Fe2+ + αKG)PHD2 at pH 6.50 with pH 8.50. The near-UV circular dichroism spectrum was virtually unchanged at elevated pH, indicating that the secondary structure did not change as a function of pH. UV–visible and Fe X-ray absorption spectroscopy indicated that the primary coordination sphere of Fe2+ changed upon increasing the pH; extended X-ray absorption fine structure analysis found a short Fe–(O/N) bond length of 1.96 Å at pH 8.50, strongly suggesting that the aquo ligand was deprotonated at this pH. Solvent isotope effects were measured during steady-sate turnover over a wide pH-range, with an inverse solvent isotope effect (SIE) of kcat observed (D2Okcat = 0.91 ± 0.03) for the acid form; a similar SIE was observed for the basic form of the enzyme (D2Okcat = 0.9 ± 0.1), with an acid equilibrium offset of ΔpKa = 0.67 ± 0.04. The inverse SIE indicated that aquo release from the active site Fe2+ immediately precedes a rate-limiting step, suggesting that turnover in this enzyme may be partially limited by the rate of O2 binding or activation, and suggesting that aquo release is relatively slow. The unusual kinetic pKa further suggested that PHD2 might function physiologically to sense both intracellular pO2 as well as pH, which could provide for feedback between anaerobic metabolism and hypoxia sensing.
Co-reporter:Shannon C. Flagg, Cristina B. Martin, Cornelius Y. Taabazuing, Breanne E. Holmes, Michael J. Knapp
Journal of Inorganic Biochemistry 2012 Volume 113() pp:25-30
Publication Date(Web):August 2012
DOI:10.1016/j.jinorgbio.2012.03.002
Two primary O2-sensors for humans are the HIF-hydroxylases, enzymes that hydroxylate specific residues of the hypoxia inducible factor-α (HIF). These enzymes are factor inhibiting HIF (FIH) and prolyl hydroxylase-2 (PHD2), each an α-ketoglutarate (αKG) dependent, non-heme Fe(II) dioxygenase. Although the two enzymes have similar active sites, FIH hydroxylates Asn803 of HIF-1α while PHD2 hydroxylates Pro402 and/or Pro564 of HIF-1α. The similar structures but unique functions of FIH and PHD2 make them prime targets for selective inhibition leading to regulatory control of diseases such as cancer and stroke. Three classes of iron chelators were tested as inhibitors for FIH and PHD2: pyridines, hydroxypyrones/hydroxypyridinones and catechols. An initial screen of the ten small molecule inhibitors at varied [αKG] revealed a non-overlapping set of inhibitors for PHD2 and FIH. Dose response curves at moderate [αKG] ([αKG] ~ KM) showed that the hydroxypyrones/hydroxypyridinones were selective inhibitors, with IC50 in the μM range, and that the catechols were generally strong inhibitors of both FIH and PHD2, with IC50 in the low μM range. As support for binding at the active site of each enzyme as the mode of inhibition, electron paramagnetic resonance (EPR) spectroscopy were used to demonstrate inhibitor binding to the metal center of each enzyme. This work shows some selective inhibition between FIH and PHD2, primarily through the use of simple aromatic or pseudo-aromatic chelators, and suggests that hydroxypyrones and hydroxypyridones may be promising chelates for FIH or PHD2 inhibition.Chelators were identified as selective inhibitors of FIH and PHD2.Highlights► Bidentate chelators were screened as inhibitors of FIH and PHD2. ► EPR of (CuII)-substituted enzymes showed active-site binding of chelators. ► Hydroxypyrones and catechols were moderately selective inhibitors.
Co-reporter:Evren Saban, Shannon C. Flagg, Michael J. Knapp
Journal of Inorganic Biochemistry 2011 Volume 105(Issue 5) pp:630-636
Publication Date(Web):May 2011
DOI:10.1016/j.jinorgbio.2011.01.007
The factor inhibiting HIF (FIH) is one of the primary oxygen sensors in human cells, controlling gene expression by hydroxylating the α-subunit of the hypoxia inducible transcription factor (HIF). As FIH is an alpha-ketoglutarate dependent non-heme iron dioxygenase, oxygen activation is thought to precede substrate hydroxylation. The coupling between oxygen activation and substrate hydroxylation was hypothesized to be very tight, in order for FIH to fulfill its function as a regulatory enzyme. Coupling was investigated by looking for reactive oxygen species production during turnover. We used alkylsulfatase (AtsK), a metabolic bacterial enzyme with a related mechanism and similar turnover frequency, for comparison, and tested both FIH and AtsK for H2O2, O2− and OH formation under steady and substrate-depleted conditions. Coupling ratios were determined by comparing the ratio of substrate consumed to product formed. We found that AtsK reacted with O2 on the seconds timescale in the absence of prime substrate, and uncoupled during turnover to produce H2O2; neither O2− nor OH were detected. In contrast, FIH was unreactive toward O2 on the minutes timescale in the absence of prime substrate, and tightly coupled during steady-state turnover; we were unable to detect any reactive oxygen species produced by FIH. We also investigated the inactivation mechanisms of these enzymes and found that AtsK likely inactivated due to deoligomerizion, whereas FIH inactivated by slow autohydroxylation. Autohydroxylated FIH could not be reactivated by dithiothreitol (DTT) nor ascorbate, suggesting that autohydroxylation is likely to be irreversible under physiological conditions.The reductive half-reaction of FIH, the factor inhibiting hypoxia inducible factor, an alpha-ketoglutarate dependent oxygenase, is tightly coupled to the oxidative half-reaction. O2 activation by FIH leads to product (R-OH) with high fidelity; FIH does not produce ROS.
Co-reporter:Evren Saban, Yuan-Han Chen, John A. Hangasky, Cornelius Y. Taabazuing, Breanne E. Holmes, and Michael J. Knapp
Biochemistry 2011 Volume 50(Issue 21) pp:
Publication Date(Web):April 1, 2011
DOI:10.1021/bi102042t
The factor inhibiting HIF (FIH) is a proximate oxygen sensor for human cells, hydroxylating Asn803 within the α-subunit of the hypoxia inducible factor (HIF). FIH is an α-ketoglutatrate (αKG)-dependent, non-heme Fe(II) dioxygenase, in which Fe(II) is coordinated by a (His2Asp) facial triad, αKG, and H2O. Hydrogen bonding among the facial triad, the HIF-Asn803 side chain, and various second-sphere residues suggests a functional role for the second coordination sphere in tuning the chemistry of the Fe(II) center. Point mutants of FIH were prepared to test the functional role of the αKG-centered (Asn205 and Asn294) or HIF-Asn803-centered (Arg238 and Gln239) second-sphere residues. The second sphere was tested for local effects on priming Fe(II) to react with O2, oxidative decarboxylation, and substrate positioning. Steady-sate kinetics were used to test for overall catalytic effects; autohydroxylation rates were used to test for priming and positioning, and electronic spectroscopy was used to assess the primary coordination sphere and the electrophilicity of αKG. Asn205 → Ala and Asn294 → Ala mutants exhibited diminished rates of steady-state turnover, while minimally affecting autohydroxylation, consistent with impaired oxidative decarboxylation. Blue-shifted metal to ligand charge transfer transitions for (Fe+αKG)FIH indicated that these point mutations destabilized the π* orbitals of αKG, further supporting a slowed rate of oxidative decarboxylation. The Arg238 → Met mutant exhibited steady-state rates too low to measure and diminished product yields, suggesting impaired substrate positioning or priming; the Arg238 → Met mutant was capable of O2 activation for the autohydroxylation reaction. The Gln239 → Asn mutant exhibited significantly slowed steady-state kinetics and diminished product yields, suggesting impaired substrate positioning or priming. As HIF binding to the Gln239 → Asn mutant stimulated autohydroxylation, it is more likely that this point mutant simply mispositions the HIF-Asn803 side chain. This work combines kinetics and spectroscopy to show that these second-sphere hydrogen bonds play roles in promoting oxidative decarboxylation, priming Fe(II) to bind O2, and positioning HIF-Asn803.
Co-reporter:Meaghan E. Germain and Michael J. Knapp
Chemical Society Reviews 2009 vol. 38(Issue 9) pp:2543-2555
Publication Date(Web):05 Jun 2009
DOI:10.1039/B809631G
The detection of chemical explosives is crucial for military and civilian safety. A confluence of chemistry and engineering continues to improve the sensitivity for several classes of explosives, and holds the promise of cheap and portable sensing. Optical and fluorescence-based sensors have been extensively researched for portable applications due to their sensitivity and portability. This tutorial review discusses chemical approaches to sensing explosives based on an optical readout, and summarizes recent advances in fluorescence strategies, including fluorescence turn-on. It is of interest to researchers working in the areas of materials chemistry or forensics.
Co-reporter:Yuan-Han Chen, Lindsay M. Comeaux, Stephen J. Eyles and Michael J. Knapp
Chemical Communications 2008 (Issue 39) pp:4768-4770
Publication Date(Web):11 Aug 2008
DOI:10.1039/B809099H
HIF-asparaginyl hydroxylase (FIH-1) normally couples O2-activation to hydroxylation of Asn803 on the α-subunit of the hypoxia-inducible factor (HIFα), a key step in pO2 sensing; in the absence of HIFα, O2-activation becomes uncoupled, leading to self-hydroxylation at Trp296 and a purple Fe(III)–O–Trp chromophore—this alternative reactivity may affect human hypoxia sensing.
Co-reporter:Meaghan E. Germain ; Thomas R. Vargo ; Beth Anne McClure ; Jeffrey J. Rack ; P. Gregory Van Patten ; Michael Odoi
Inorganic Chemistry 2008 Volume 47(Issue 14) pp:6203-6211
Publication Date(Web):June 25, 2008
DOI:10.1021/ic702469q
Nitroaromatics and nitroalkanes quench the fluorescence of Zn(Salophen) (H2Salophen = N,N′-phenylene-bis-(3,5-di-tert-butylsalicylideneimine); ZnLR) complexes. A structurally related family of ZnLR complexes (R = OMe, di-tBu, tBu, Cl, NO2) were prepared, and the mechanisms of fluorescence quenching by nitroaromatics were studied by a combined kinetics and spectroscopic approach. The fluorescent quantum yields for ZnLR were generally high (Φ ∼ 0.3) with sub-nanosecond fluorescence lifetimes. The fluorescence of ZnLR was quenched by nitroaromatic compounds by a mixture of static and dynamic pathways, reflecting the ZnLR ligand bulk and reduction potential. Steady-state Stern-Volmer plots were curved for ZnLR with less-bulky substituents (R = OMe, NO2), suggesting that both static and dynamic pathways were important for quenching. Transient Stern-Volmer data indicated that the dynamic pathway dominated quenching for ZnLR with bulky substituents (R = tBu, DtBu). The quenching rate constants with varied nitroaromatics (ArNO2) followed the driving force dependence predicted for bimolecular electron transfer: ZnL* + ArNO2 → ZnL+ + ArNO2−. A treatment of the diffusion-corrected quenching rates with Marcus theory yielded a modest reorganization energy (λ = 25 kcal/mol), and a small self-exchange reorganization energy for ZnL*/ZnL+ (ca. 20 kcal/mol) was estimated from the Marcus cross-relation, suggesting that metal phenoxyls may be robust biological redox cofactors. Electronic structure calculations indicated very small changes in bond distances for the ZnL → ZnL+ oxidation, suggesting that solvation was the dominant contributor to the observed reorganization energy. These mechanistic insights provide information that will be helpful to further develop ZnLR as sensors, as well as for potential photoinduced charge transfer chemistry.
Co-reporter:Halil Bayraktar, Sudhanshu Srivastava, Chang-Cheng You, Vincent M. Rotello and Michael J. Knapp
Soft Matter 2008 vol. 4(Issue 4) pp:751-756
Publication Date(Web):24 Jan 2008
DOI:10.1039/B716386J
Selective surface recognition by proteins provides programmed bottom-up assembly of synthetic nanomaterials. We have investigated the controlled self-assembly of functionalized gold nanoparticles (Au-TAsp) with cytochrome c (Cyt c) and apoCyt c through complementary electrostatic interactions. Au-TAsp formed discrete, water-soluble adducts with native Cyt c, whereas unfolded apoCyt c induced nanocomposite formation at high Cyt c : Au-TAsp ratios. The binding of random-coil apoCyt c to Au-TAsp at low ratios induced α-helix formation in soluble nanocomposites, but at elevated ratios insoluble micron-scale aggregates were formed. The local structure of the assemblies was critically dependent on the Cyt c : Au-TAsp ratio. The dispersibility of apoCyt c–Au-TAsp was pH dependent, providing rapid and reversible control over nanocomposite assembly. The apoCyt c–Au-TAsp aggregates could likewise be disassembled through proteolytic cleavage of apoCyt c, demonstrating the ability to selectively remodel these hybrid materials.
Co-reporter:Yuan-Han Chen, Lindsay M. Comeaux, Robert W. Herbst, Evren Saban, David C. Kennedy, Michael J. Maroney, Michael J. Knapp
Journal of Inorganic Biochemistry 2008 Volume 102(Issue 12) pp:2120-2129
Publication Date(Web):December 2008
DOI:10.1016/j.jinorgbio.2008.07.018
Hypoxia sensing is the generic term for pO2-sensing in humans and other higher organisms. These cellular responses to pO2 are largely controlled by enzymes that belong to the Fe(II) α-ketoglutarate (αKG) dependent dioxygenase superfamily, including the human enzyme called the factor inhibiting HIF (FIH-1), which couples O2-activation to the hydroxylation of the hypoxia inducible factor α (HIFα). Uncoupled O2-activation by human FIH-1 was studied by exposing the resting form of FIH-1 (αKG + Fe)FIH-1, to air in the absence of HIFα. Uncoupling lead to two distinct enzyme oxidations, one a purple chromophore (λmax = 583 nm) arising from enzyme auto-hydroxylation of Trp296, forming an Fe(III)–O–Trp296 chromophore [Y.-H. Chen, L.M. Comeaux, S.J. Eyles, M.J. Knapp, Chem. Commun. (2008), doi:10.1039/B809099H]; the other a yellow chromophore due to Fe(III) in the active site, which under some conditions also contained variable levels of an oxygenated surface residue (oxo)Met275. The kinetics of purple FIH-1 formation were independent of Fe(II) and αKG concentrations, however, product yield was saturable with increasing [αKG] and required excess Fe(II). Yellow FIH-1 was formed from (succinate + Fe)FIH-1, or by glycerol addition to (αKG + Fe)FIH-1, suggesting that glycerol could intercept the active oxidant from the FIH-1 active site and prevent hydroxylation. Both purple and yellow FIH-1 contained high-spin, rhombic Fe(III) centers, as shown by low temperature EPR. XAS indicated distorted octahedral Fe(III) geometries, with subtle differences in inner-shell ligands for yellow and purple FIH-1. EPR of Co(II)-substituted FIH-1 (αKG + Co)FIH-1, indicated a mixture of 5-coordinate and 6-coordinate enzyme forms, suggesting that resting FIH-1 can readily undergo uncoupled O2-activation by loss of an H2O ligand from the metal center.
Co-reporter:Halil Bayraktar, Partha S. Ghosh, Vincent M. Rotello and Michael J. Knapp
Chemical Communications 2006 (Issue 13) pp:1390-1392
Publication Date(Web):21 Feb 2006
DOI:10.1039/B516096K
Functionalized gold nanoparticles bind selectively to cytochrome c or cytochrome c peroxidase and inhibit enzyme turnover.
Co-reporter:Meaghan E. Germain
Inorganic Chemistry () pp:
Publication Date(Web):October 2, 2008
DOI:10.1021/ic801317x
Peroxide-based explosives, like triacetone triperoxide (TATP), are important targets for detection because of their broad use in improvised explosives but pose challenges. We report a highly sensitive turn-on fluorescence detection for H2O2 and organic peroxides, including TATP. The detection strategy relies on oxidative deboronation to unmask H2Salen, which subsequently binds Zn2+ to form fluorescent Zn(Salen). Sensitivity is excellent, with detection limits below 10 nM for H2O2, TATP, and benzoyl peroxide. In addition, acid treatment is necessary to sense TATP, suggesting the potential to discriminate between H2O2 and TATP based upon minimal sample pretreatment.
Co-reporter:Yuan-Han Chen, Lindsay M. Comeaux, Stephen J. Eyles and Michael J. Knapp
Chemical Communications 2008(Issue 39) pp:NaN4770-4770
Publication Date(Web):2008/08/11
DOI:10.1039/B809099H
HIF-asparaginyl hydroxylase (FIH-1) normally couples O2-activation to hydroxylation of Asn803 on the α-subunit of the hypoxia-inducible factor (HIFα), a key step in pO2 sensing; in the absence of HIFα, O2-activation becomes uncoupled, leading to self-hydroxylation at Trp296 and a purple Fe(III)–O–Trp chromophore—this alternative reactivity may affect human hypoxia sensing.
Co-reporter:Meaghan E. Germain and Michael J. Knapp
Chemical Society Reviews 2009 - vol. 38(Issue 9) pp:NaN2555-2555
Publication Date(Web):2009/06/05
DOI:10.1039/B809631G
The detection of chemical explosives is crucial for military and civilian safety. A confluence of chemistry and engineering continues to improve the sensitivity for several classes of explosives, and holds the promise of cheap and portable sensing. Optical and fluorescence-based sensors have been extensively researched for portable applications due to their sensitivity and portability. This tutorial review discusses chemical approaches to sensing explosives based on an optical readout, and summarizes recent advances in fluorescence strategies, including fluorescence turn-on. It is of interest to researchers working in the areas of materials chemistry or forensics.
.jpg)