Co-reporter:Gwendolyn Moise, Nathan M. Gallup, Anastassia N. Alexandrova, Alvan C. Hengge, and Sean J. Johnson
Biochemistry 2015 Volume 54(Issue 42) pp:6490-6500
Publication Date(Web):October 7, 2015
DOI:10.1021/acs.biochem.5b00496
Catalysis in protein tyrosine phosphatases (PTPs) involves movement of a protein loop called the WPD loop that brings a conserved aspartic acid into the active site to function as a general acid. Mutation of the tryptophan in the WPD loop of the PTP YopH to any other residue with a planar, aromatic side chain (phenylalanine, tyrosine, or histidine) disables general acid catalysis. Crystal structures reveal these conservative mutations leave this critical loop in a catalytically unproductive, quasi-open position. Although the loop positions in crystal structures are similar for all three conservative mutants, the reasons inhibiting normal loop closure differ for each mutant. In the W354F and W354Y mutants, steric clashes result from six-membered rings occupying the position of the five-membered ring of the native indole side chain. The histidine mutant dysfunction results from new hydrogen bonds stabilizing the unproductive position. The results demonstrate how even modest modifications can disrupt catalytically important protein dynamics. Crystallization of all the catalytically compromised mutants in the presence of vanadate gave rise to vanadate dimers at the active site. In W354Y and W354H, a divanadate ester with glycerol is observed. Such species have precedence in solution and are known from the small molecule crystal database. Such species have not been observed in the active site of a phosphatase, as a functional phosphatase would rapidly catalyze their decomposition. The compromised functionality of the mutants allows the trapping of species that undoubtedly form in solution and are capable of binding at the active sites of PTPs, and, presumably, other phosphatases. In addition to monomeric vanadate, such higher-order vanadium-based molecules are likely involved in the interaction of vanadate with PTPs in solution.
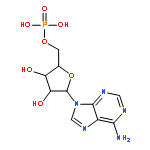
Co-reporter:Vyacheslav I. Kuznetsov, Alvan C. Hengge, and Sean J. Johnson
Biochemistry 2012 Volume 51(Issue 49) pp:
Publication Date(Web):November 12, 2012
DOI:10.1021/bi300908y
The recently discovered 150-residue human VHZ (VH1-related protein, Z member) is one of the smallest protein tyrosine phosphatases (PTPs) known and contains only the minimal structural elements common to all PTPs. We report a substrate screening analysis and a crystal structure of the VHZ complex with vanadate at 1.1 Å resolution, with a detailed structural comparison with other members of the protein tyrosine phosphatase family, including classical tyrosine-specific protein tyrosine phosphatases (PTPs) and dual-specificity phosphatases (DSPs). A screen with 360 phosphorylated peptides shows VHZ efficiently catalyzes the hydrolysis of phosphotyrosine (pY)-containing peptides but exhibits no activity toward phosphoserine (pS) or phosphothreonine (pT) peptides. The new structure reveals a deep and narrow active site more typical of the classical tyrosine-specific PTPs. Despite the high degrees of structural and sequence similarity between VHZ and classical PTPs, its general acid IPD-loop is most likely conformationally rigid, in contrast to the flexible WPD counterpart of classical PTPs. VHZ also lacks substrate recognition domains and other domains typically found on classical PTPs. It is therefore proposed that VHZ is more properly classified as an atypical PTP rather than an atypical DSP, as has been suggested.
Co-reporter:Jeremy W. Bakelar, Dariusz A. Sliwa, Sean J. Johnson
Archives of Biochemistry and Biophysics (May 2013) Volume 533(Issues 1–2) pp:62-68
Publication Date(Web):May 2013
DOI:10.1016/j.abb.2013.02.017
Co-reporter:Tiago A.S. Brandão, Sean J. Johnson, Alvan C. Hengge
Archives of Biochemistry and Biophysics (1 September 2012) Volume 525(Issue 1) pp:53-59
Publication Date(Web):1 September 2012
DOI:10.1016/j.abb.2012.06.002
.jpg)




![Ethanesulfonic acid, 2-[[(2S)-2-hydroxypropyl]thio]-](http://img.cochemist.com/ccimg/244400/244301-87-9.png)
![Ethanesulfonic acid, 2-[[(2S)-2-hydroxypropyl]thio]-](http://img.cochemist.com/ccimg/244400/244301-87-9_b.png)


![L-Ornithine,N5-[(methylamino)(methylimino)methyl]-](http://img.cochemist.com/ccimg/30400/30344-00-4.png)
![L-Ornithine,N5-[(methylamino)(methylimino)methyl]-](http://img.cochemist.com/ccimg/30400/30344-00-4_b.png)
![L-Ornithine,N5-[(dimethylamino)iminomethyl]-](http://img.cochemist.com/ccimg/30400/30315-93-6.png)
![L-Ornithine,N5-[(dimethylamino)iminomethyl]-](http://img.cochemist.com/ccimg/30400/30315-93-6_b.png)