Co-reporter:Xinying Shi and Simpson Joseph
Biochemistry 2016 Volume 55(Issue 45) pp:6344
Publication Date(Web):October 25, 2016
DOI:10.1021/acs.biochem.6b00921
Release factors 1 and 2 (RF1 and RF2, respectively) bind to ribosomes that have a stop codon in the A site and catalyze the release of the newly synthesized protein. Following peptide release, the dissociation of RF1 and RF2 from the ribosome is accelerated by release factor 3 (RF3). The mechanism for RF3-promoted dissociation of RF1 and RF2 is unclear. It was previously proposed that RF3 hydrolyzes GTP and dissociates from the ribosome after RF1 dissociation. Here we monitored directly the dissociation kinetics of RF1 and RF3 using Förster resonance energy transfer-based assays. In contrast to the previous model, our data show that RF3 hydrolyzes GTP and dissociates from the ribosome before RF1 dissociation. We propose that RF3 stabilizes the ratcheted state of the ribosome, which consequently accelerates the dissociation of RF1 and RF2.
Co-reporter:Bhubanananda Sahu, Prashant K. Khade, and Simpson Joseph
Biochemistry 2013 Volume 52(Issue 38) pp:
Publication Date(Web):August 30, 2013
DOI:10.1021/bi4008879
Accurate decoding of mRNA requires the precise interaction of protein factors and tRNAs with the ribosome. X-ray crystallography and cryo-electron microscopy have provided detailed structural information about the 70S ribosome with protein factors and tRNAs trapped during translation. Crystal structures showed that one of the universally conserved 16S rRNA bases, A55, in the shoulder domain of the 30S subunit interacts with elongation factors Tu and G (EF-Tu and EF-G, respectively). The exact functional role of A55 in protein synthesis is not clear. We changed A55 to U and analyzed the effect of the mutation on the elongation cycle of protein synthesis using functional assays. Expression of 16S rRNA with the A55U mutation in cells confers a dominant lethal phenotype. Additionally, ribosomes with the A55U mutation in 16S rRNA show substantially reduced in vitro protein synthesis activity. Equilibrium binding studies showed that the A55U mutation considerably inhibited the binding of the EF-Tu·GTP·tRNA ternary complex to the ribosome. Furthermore, the A55U mutation slightly inhibited the peptidyl transferase reaction, the binding of EF-G·GTP to the ribosome, and mRNA–tRNA translocation. These results indicate that A55 is important for fine-tuning the activity of the ribosome during the elongation cycle of protein synthesis.
Co-reporter:Bhubanananda Sahu, Prashant K. Khade, and Simpson Joseph
Biochemistry 2012 Volume 51(Issue 38) pp:
Publication Date(Web):August 31, 2012
DOI:10.1021/bi300930r
Ribosomes are RNA–protein complexes responsible for protein synthesis. A dominant structural motif in the rRNAs is an RNA helix capped with a four-nucleotide loop, called a tetraloop. The sequence of the tetraloop is invariant at some positions in the rRNAs but is highly variable at other positions. The biological reason for the conservation of the tetraloop sequence at specific positions in the rRNAs is not clear. In the 16S rRNA, the GAAA tetraloop in helix 8 and the UACG tetraloop in helix 14 are highly conserved and located near the binding site for EF-Tu and EF-G. To investigate whether the structural stability of the tetraloop or the precise sequence of the tetraloop is important for function, we separately changed the GAAA tetraloop in helix 8 to a UACG tetraloop and the UACG tetraloop in helix 14 to a GAAA tetraloop. The effects of the tetraloop replacements on protein synthesis were analyzed in vivo and in vitro. Replacement of the tetraloops in helices 8 and 14 did not significantly affect the growth rate of the Escherichia coli (Δ7rrn) strain. However, the mutant ribosomes showed a slightly reduced rate of protein synthesis in vitro. In addition, we observed a 2-fold increase in the error rate of translation with the mutant ribosomes, which is consistent with an earlier report. Our results suggest that the tetraloops in helices 8 and 14 are highly conserved mainly for their structural stability and the precise sequences of these tetraloops are not critical for protein synthesis.
Co-reporter:Andrew Field, Byron Hetrick, Merrill Mathew, and Simpson Joseph
Biochemistry 2010 Volume 49(Issue 43) pp:
Publication Date(Web):September 28, 2010
DOI:10.1021/bi1012047
Class I peptide release factors 1 and 2 (RF1 and RF2, respectively) recognize the stop codons in the ribosomal decoding center and catalyze peptidyl-tRNA hydrolysis. High-fidelity stop codon recognition by these release factors is essential for accurate peptide synthesis and ribosome recycling. X-ray crystal structures of RF1 and RF2 bound to the ribosome have identified residues in the mRNA−protein interface that appear to be critical for stop codon recognition. Especially interesting is a conserved histidine in all bacterial class I release factors that forms a stacking interaction with the second base of the stop codon. Here we analyzed the functional significance of this conserved histidine (position 197 in Escherichia coli) of RF1 by point mutagenesis to alanine. Equilibrium binding studies and transient-state kinetic analysis have shown that the histidine is essential for binding with high affinity to the ribosome. Furthermore, analysis of the binding data indicates a conformational change within the RF1·ribosome complex that results in a more tightly bound state. The rate of peptidyl-tRNA hydrolysis was also reduced significantly, more than the binding data would suggest, implying a defect in the orientation of the GGQ domain without the histidine residue.
Co-reporter:Byron Hetrick, Prashant K. Khade, Kristin Lee, Jenise Stephen, Alex Thomas and Simpson Joseph
Biochemistry 2010 Volume 49(Issue 33) pp:
Publication Date(Web):July 20, 2010
DOI:10.1021/bi1009776
The selection of aminoacyl-tRNAs by the ribosome is a fundamental step in the elongation cycle of protein synthesis. tRNA selection is a multistep process that ensures only correct aminoacyl-tRNAs are accepted while incorrect aminoacyl-tRNAs are rejected. A key step in tRNA selection is the formation of base pairs between the anticodon of the aminoacyl-tRNA and the mRNA codon in the A site, called “codon recognition”. Here, we report the development of a new, fluorescence-based, kinetic assay for monitoring codon recognition by the ribosome. Using this assay, we show that codon recognition is a second-order binding step under optimal conditions. Additionally, we show that at low Mg2+ concentrations, the polyamines spermine and spermidine stimulate codon recognition by the ribosome without a loss of fidelity. Polyamines may accelerate codon recognition by altering the structure and dynamics of the anticodon arm of the aminoacyl-tRNA.
Co-reporter:Byron Hetrick, Kristin Lee and Simpson Joseph
Biochemistry 2009 Volume 48(Issue 47) pp:
Publication Date(Web):October 29, 2009
DOI:10.1021/bi901577d
Recognition of stop codons by class I release factors is a fundamental step in the termination phase of protein synthesis. Since premature termination is costly to the cell, release factors have to efficiently discriminate between stop and sense codons. To understand the mechanism of discrimination between stop and sense codons, we developed a new, pre-steady state kinetic assay to monitor the interaction of RF1 with the ribosome. Our results show that RF1 associates with similar association rate constants with ribosomes programmed with stop or sense codons. However, dissociation of RF1 from sense codons is as much as 3 orders of magnitude faster than from stop codons. Interestingly, the affinity of RF1 for ribosomes programmed with different sense codons does not correlate with the defects in peptide release. Thus, discrimination against sense codons is achieved with both an increase in the dissociation rates and a decrease in the rate of peptide release. These results suggest that sense codons inhibit conformational changes necessary for RF1 to stably bind to the ribosome and catalyze peptide release.
Co-reporter:Eileen Chen, Simpson Joseph
Biochimie (July 2015) Volume 114() pp:147-154
Publication Date(Web):July 2015
DOI:10.1016/j.biochi.2015.02.005
Co-reporter:Krista Trappl, Simpson Joseph
Journal of Molecular Biology (27 March 2016) Volume 428(Issue 6) pp:1333-1344
Publication Date(Web):27 March 2016
DOI:10.1016/j.jmb.2016.01.021
•Transition metal ion fluorescence resonance energy transfer analysis shows that RF1 free in solution is in a closed conformation.•Binding to the ribosome induces the open conformation of RF1.•RF1 and RF2 may use an induced-fit mechanism to recognize stop codons.Bacterial translation termination is triggered when a stop codon arrives at the ribosomal A site. Stop codons are recognized by class I release factors (RF1 and RF2 in Escherichia coli), which bind to the ribosome and catalyze the release of the newly synthesized protein. Crystal structures showed that RF1 and RF2 are in an open conformation when bound to the ribosome but are in a closed conformation when not bound to the ribosome. It is not clear whether only the open form of RF1 and RF2 binds to the ribosome. Alternatively, the closed form of RF1 and RF2 may bind to the ribosome and undergo a conformational change to the open state upon binding. We used transition metal ion fluorescence resonance energy transfer experiments to monitor precisely the conformation of RF1 in the absence and presence of the ribosome. Our results indicate that RF1 undergoes a large conformational change from a closed to an open form upon binding to the ribosome. Our results are consistent with the mechanism, in which high termination fidelity is achieved by linking stop codon recognition by RF1 to the change in conformation from closed to open state.Download high-res image (253KB)Download full-size image
Co-reporter:Prashant K. Khade, Xinying Shi, Simpson Joseph
Journal of Molecular Biology (23 October 2013) Volume 425(Issue 20) pp:3778-3789
Publication Date(Web):23 October 2013
DOI:10.1016/j.jmb.2013.02.038
► Mechanism of tRNA selection is analyzed using modified mRNAs. ► 2′-Deoxynucleotide substitutions in the mRNA inhibited key steps in tRNA selection. ► 2′-Fluoro substitutions in the mRNA have only modest effects on tRNA selection. ► Shape complementarity in the decoding center is critical for tRNA selection.Accurate tRNA selection by the ribosome is essential for the synthesis of functional proteins. Previous structural studies indicated that the ribosome distinguishes between cognate and near-cognate tRNAs by monitoring the geometry of the codon–anticodon helix in the decoding center using the universally conserved 16S ribosomal RNA bases G530, A1492 and A1493. These bases form hydrogen bonds with the 2′-hydroxyl groups of the codon–anticodon helix, which are expected to be disrupted with a near-cognate codon–anticodon helix. However, a recent structural study showed that G530, A1492 and A1493 form hydrogen bonds in a manner identical with that of both cognate and near-cognate codon–anticodon helices. To understand how the ribosome discriminates between cognate and near-cognate tRNAs, we made 2′-deoxynucleotide and 2′-fluoro substituted mRNAs, which disrupt the hydrogen bonds between the A site codon and G530, A1492 and A1493. Our results show that multiple 2′-deoxynucleotide substitutions in the mRNA substantially inhibit tRNA selection, whereas multiple 2′-fluoro substitutions in the mRNA have only modest effects on tRNA selection. Furthermore, the miscoding antibiotics paromomycin and streptomycin rescue the defects in tRNA selection with the multiple 2′-deoxynucleotide substituted mRNA. These results suggest that steric complementarity in the decoding center is more important than the hydrogen bonds between the A site codon and G530, A1492 and A1493 for tRNA selection.Download high-res image (405KB)Download full-size image
.jpg)






















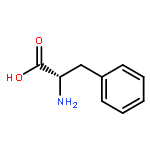
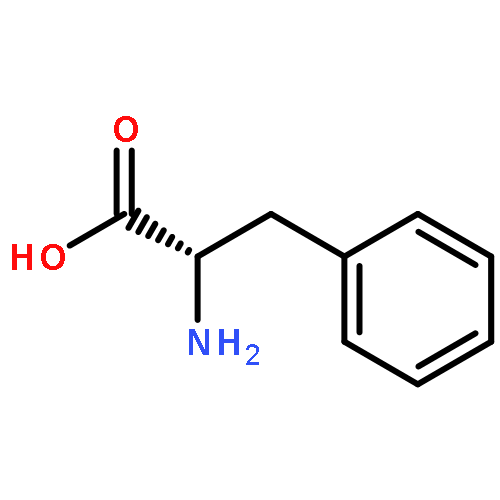
![2-Propenamide,N-[(1S)-1-(hydroxymethyl)-2-[(R)-[(methylthio)methyl]sulfinyl]ethyl]-3-(1,2,3,4-tetrahydro-6-methyl-2,4-dioxo-5-pyrimidinyl)-,(2E)-](http://img.cochemist.com/ccimg/1500/1404-64-4.png)
![2-Propenamide,N-[(1S)-1-(hydroxymethyl)-2-[(R)-[(methylthio)methyl]sulfinyl]ethyl]-3-(1,2,3,4-tetrahydro-6-methyl-2,4-dioxo-5-pyrimidinyl)-,(2E)-](http://img.cochemist.com/ccimg/1500/1404-64-4_b.png)