Co-reporter:David Oyen, R. Bryn Fenwick, Phillip C. Aoto, Robyn L. Stanfield, Ian A. Wilson, H. Jane Dyson, and Peter E. Wright
Journal of the American Chemical Society August 16, 2017 Volume 139(Issue 32) pp:11233-11233
Publication Date(Web):July 24, 2017
DOI:10.1021/jacs.7b05958
The rate-determining step in the catalytic cycle of E. coli dihydrofolate reductase is tetrahydrofolate (THF) product release, which can occur via an allosteric or an intrinsic pathway. The allosteric pathway, which becomes accessible when the reduced cofactor NADPH is bound, involves transient sampling of a higher energy conformational state, greatly increasing the product dissociation rate as compared to the intrinsic pathway that obtains when NADPH is absent. Although the kinetics of this process are known, the enzyme structure and the THF product conformation in the transiently formed excited state remain elusive. Here, we use side-chain proton NMR relaxation dispersion measurements, X-ray crystallography, and structure-based chemical shift predictions to explore the structural basis of allosteric product release. In the excited state of the E:THF:NADPH product release complex, the reduced nicotinamide ring of the cofactor transiently enters the active site where it displaces the pterin ring of the THF product. The p-aminobenzoyl-l-glutamate tail of THF remains weakly bound in a widened binding cleft. Thus, through transient entry of the nicotinamide ring into the active site, the NADPH cofactor remodels the enzyme structure and the conformation of the THF to form a weakly populated excited state that is poised for rapid product release.
Co-reporter:R. Bryn Fenwick, David Oyen and Peter E. Wright
Physical Chemistry Chemical Physics 2016 vol. 18(Issue 8) pp:5789-5798
Publication Date(Web):24 Sep 2015
DOI:10.1039/C5CP04670J
Carr–Purcell–Meiboom–Gill (CPMG) relaxation dispersion measurements are a valuable tool for the characterization of structural transitions on the micro–millisecond timescale. While the measurement of 15N relaxation dispersion is now routine, the measurements with alternative nuclei remain limited. Here we report 15N as well as 1H R2 relaxation dispersion measurements of the N23PP/S148A “dynamic knockout” mutant of dihydrofolate reductase. The 1H dispersion measurements are complementary to 15N data as many additional residues are observed to have dispersive behavior for the 1H nucleus. Simultaneous fitting of the dispersion profiles for the two nuclei increases the accuracy of exchange parameters determined for individual residues and clustered groups of residues. The different sensitivity of the two nuclei to changes in backbone torsional angles, ring currents, and hydrogen bonding effects provides important insights into the nature of the structural changes that take place during the exchange process. We observe clear evidence of direct and indirect hydrogen bond effects for the 15N and 1H chemical shift changes in the active-site, modulation of ring current shielding in the CD-loop and backbone torsional changes in a cluster of residues associated with the C-terminus. This work demonstrates the power of combined 1H and 15N probes for the study of backbone dynamics on the micro-millisecond timescale though the analysis of chemical shift changes.
Co-reporter:Josephine C. Ferreon;Alexander S. Krois;H. Jane Dyson;Maria A. Martinez-Yamout
PNAS 2016 Volume 113 (Issue 13 ) pp:E1853-E1862
Publication Date(Web):2016-03-29
DOI:10.1073/pnas.1602487113
An important component of the activity of p53 as a tumor suppressor is its interaction with the transcriptional coactivators
cyclic-AMP response element-binding protein (CREB)-binding protein (CBP) and p300, which activate transcription of p53-regulated
stress response genes and stabilize p53 against ubiquitin-mediated degradation. The highest affinity interactions are between
the intrinsically disordered N-terminal transactivation domain (TAD) of p53 and the TAZ1 and TAZ2 domains of CBP/p300. The
NMR spectra of simple binary complexes of the TAZ1 and TAZ2 domains with the p53TAD suffer from exchange broadening, but innovations
in construct design and isotopic labeling have enabled us to obtain high-resolution structures using fusion proteins, uniformly
labeled in the case of the TAZ2–p53TAD fusion and segmentally labeled through transintein splicing for the TAZ1–p53TAD fusion.
The p53TAD is bipartite, with two interaction motifs, termed AD1 and AD2, which fold to form short amphipathic helices upon
binding to TAZ1 and TAZ2 whereas intervening regions of the p53TAD remain flexible. Both the AD1 and AD2 motifs bind to hydrophobic
surfaces of the TAZ domains, with AD2 making more extensive hydrophobic contacts consistent with its greater contribution
to the binding affinity. Binding of AD1 and AD2 is synergistic, and structural studies performed with isolated motifs can
be misleading. The present structures of the full-length p53TAD complexes demonstrate the versatility of the interactions
available to an intrinsically disordered domain containing bipartite interaction motifs and provide valuable insights into
the structural basis of the affinity changes that occur upon stress-related posttranslational modification.
Co-reporter:Shu-ou Shan;Chris Chi;Camille Z. McAvoy;Fu-Cheng Liang;Gerard Kroon
PNAS 2016 Volume 113 (Issue 12 ) pp:E1615-E1624
Publication Date(Web):2016-03-22
DOI:10.1073/pnas.1524777113
Membrane protein biogenesis poses enormous challenges to cellular protein homeostasis and requires effective molecular chaperones.
Compared with chaperones that promote soluble protein folding, membrane protein chaperones require tight spatiotemporal coordination
of their substrate binding and release cycles. Here we define the chaperone cycle for cpSRP43, which protects the largest
family of membrane proteins, the light harvesting chlorophyll a/b-binding proteins (LHCPs), during their delivery. Biochemical
and NMR analyses demonstrate that cpSRP43 samples three distinct conformations. The stromal factor cpSRP54 drives cpSRP43
to the active state, allowing it to tightly bind substrate in the aqueous compartment. Bidentate interactions with the Alb3
translocase drive cpSRP43 to a partially inactive state, triggering selective release of LHCP’s transmembrane domains in a
productive unloading complex at the membrane. Our work demonstrates how the intrinsic conformational dynamics of a chaperone
enables spatially coordinated substrate capture and release, which may be general to other ATP-independent chaperone systems.
Co-reporter:David Oyen; R. Bryn Fenwick; Robyn L. Stanfield; H. Jane Dyson
Journal of the American Chemical Society 2015 Volume 137(Issue 29) pp:9459-9468
Publication Date(Web):July 6, 2015
DOI:10.1021/jacs.5b05707
The enzyme dihydrofolate reductase (DHFR, E) from Escherichia coli is a paradigm for the role of protein dynamics in enzyme catalysis. Previous studies have shown that the enzyme progresses through the kinetic cycle by modulating the dynamic conformational landscape in the presence of substrate dihydrofolate (DHF), product tetrahydrofolate (THF), and cofactor (NADPH or NADP+). This study focuses on the quantitative description of the relationship between protein fluctuations and product release, the rate-limiting step of DHFR catalysis. NMR relaxation dispersion measurements of millisecond time scale motions for the E:THF:NADP+ and E:THF:NADPH complexes of wild-type and the Leu28Phe (L28F) point mutant reveal conformational exchange between an occluded ground state and a low population of a closed state. The backbone structures of the occluded ground states of the wild-type and mutant proteins are very similar, but the rates of exchange with the closed excited states are very different. Integrated analysis of relaxation dispersion data and THF dissociation rates measured by stopped-flow spectroscopy shows that product release can occur by two pathways. The intrinsic pathway consists of spontaneous product dissociation and occurs for all THF-bound complexes of DHFR. The allosteric pathway features cofactor-assisted product release from the closed excited state and is utilized only in the E:THF:NADPH complexes. The L28F mutation alters the partitioning between the pathways and results in increased flux through the intrinsic pathway relative to the wild-type enzyme. This repartitioning could represent a general mechanism to explain changes in product release rates in other E. coli DHFR mutants.
Co-reporter:Munehito Arai;Kenji Sugase;H. Jane Dyson
PNAS 2015 Volume 112 (Issue 31 ) pp:9614-9619
Publication Date(Web):2015-08-04
DOI:10.1073/pnas.1512799112
Intrinsically disordered proteins (IDPs) frequently function in protein interaction networks that regulate crucial cellular
signaling pathways. Many IDPs undergo transitions from disordered conformational ensembles to folded structures upon binding
to their cellular targets. Several possible binding mechanisms for coupled folding and binding have been identified: folding
of the IDP after association with the target (“induced fit”), or binding of a prefolded state in the conformational ensemble
of the IDP to the target protein (“conformational selection”), or some combination of these two extremes. The interaction
of the intrinsically disordered phosphorylated kinase-inducible domain (pKID) of the cAMP-response element binding (CREB)
protein with the KIX domain of a general transcriptional coactivator CREB-binding protein (CBP) provides an example of the
induced-fit mechanism. Here we show by NMR relaxation dispersion experiments that a different intrinsically disordered ligand,
the transactivation domain of the transcription factor c-Myb, interacts with KIX at the same site as pKID but via a different
binding mechanism that involves elements of conformational selection and induced fit. In contrast to pKID, the c-Myb activation
domain has a strong propensity for spontaneous helix formation in its N-terminal region, which binds to KIX in a predominantly
folded conformation. The C-terminal region of c-Myb exhibits a much smaller helical propensity and likely folds via an induced-fit
process after binding to KIX. We propose that the intrinsic secondary structure propensities of pKID and c-Myb determine their
binding mechanisms, consistent with their functions as inducible and constitutive transcriptional activators.
Co-reporter:Russell G. Burge, Maria A. Martinez-Yamout, H. Jane Dyson, and Peter E. Wright
Biochemistry 2014 Volume 53(Issue 9) pp:
Publication Date(Web):February 13, 2014
DOI:10.1021/bi401675h
The interactions of the human double-stranded RNA-binding zinc finger protein JAZ with RNA or DNA were investigated using electrophoretic mobility-shift assays, isothermal calorimetry, and nuclear magnetic resonance spectroscopy. Consistent with previous reports, JAZ has very low affinity for duplex DNA or single-stranded RNA, but it binds preferentially to double-stranded RNA (dsRNA) with no detectable sequence specificity. The affinity of JAZ for dsRNA is unaffected by local structural features such as loops, overhangs, and bulges, provided a sufficient length of reasonably well-structured A-form RNA (about 18 bp for a single zinc finger) is present. Full-length JAZ contains four Cys2His2 zinc fingers (ZF1–4) and has the highest apparent affinity for dsRNA; two-finger constructs ZF12 and ZF23 have lower affinity, and ZF34 binds even more weakly. The fourth zinc finger, ZF4, has no measurable RNA-binding affinity. Single zinc finger constructs ZF1, ZF2, and ZF3 show evidence for multiple-site binding on the minimal RNA. Fitting of quantitative NMR titration and isothermal calorimetry data to a two-site binding model gave Kd1 ∼ 10 μM and Kd2 ∼ 100 μM. Models of JAZ–RNA complexes were generated using the high-ambiguity-driven biomolecular docking (HADDOCK) program. Single zinc fingers bind to the RNA backbone without sequence specificity, forming complexes with contacts between the RNA minor groove and residues in the N-terminal β strands and between the major groove and residues in the helix–kink–helix motif. We propose that the non-sequence-specific interaction between the zinc fingers of JAZ with dsRNA is dependent only on the overall shape of the A-form RNA.
Co-reporter:Lisa M. Tuttle, H. Jane Dyson, and Peter E. Wright
Biochemistry 2014 Volume 53(Issue 7) pp:
Publication Date(Web):February 5, 2014
DOI:10.1021/bi4015314
The three-dimensional structures of the dihydrofolate reductase enzymes from Escherichia coli (ecDHFR or ecE) and Homo sapiens (hDHFR or hE) are very similar, despite a rather low level of sequence identity. Whereas the active site loops of ecDHFR undergo major conformational rearrangements during progression through the reaction cycle, hDHFR remains fixed in a closed loop conformation in all of its catalytic intermediates. To elucidate the structural and dynamic differences between the human and E. coli enzymes, we conducted a comprehensive analysis of side chain flexibility and dynamics in complexes of hDHFR that represent intermediates in the major catalytic cycle. Nuclear magnetic resonance relaxation dispersion experiments show that, in marked contrast to the functionally important motions that feature prominently in the catalytic intermediates of ecDHFR, millisecond time scale fluctuations cannot be detected for hDHFR side chains. Ligand flux in hDHFR is thought to be mediated by conformational changes between a hinge-open state when the substrate/product-binding pocket is vacant and a hinge-closed state when this pocket is occupied. Comparison of X-ray structures of hinge-open and hinge-closed states shows that helix αF changes position by sliding between the two states. Analysis of χ1 rotamer populations derived from measurements of 3JCγCO and 3JCγN couplings indicates that many of the side chains that contact helix αF exhibit rotamer averaging that may facilitate the conformational change. The χ1 rotamer adopted by the Phe31 side chain depends upon whether the active site contains the substrate or product. In the holoenzyme (the binary complex of hDHFR with reduced nicotinamide adenine dinucleotide phosphate), a combination of hinge opening and a change in the Phe31 χ1 rotamer opens the active site to facilitate entry of the substrate. Overall, the data suggest that, unlike ecDHFR, hDHFR requires minimal backbone conformational rearrangement as it proceeds through its enzymatic cycle, but that ligand flux is brokered by more subtle conformational changes that depend on the side chain motions of critical residues.
Co-reporter:Phillip C. Aoto, Chiaki Nishimura, H. Jane Dyson, and Peter E. Wright
Biochemistry 2014 Volume 53(Issue 23) pp:
Publication Date(Web):May 23, 2014
DOI:10.1021/bi500478m
Apomyoglobin folds via sequential helical intermediates that are formed by rapid collapse of the A, B, G, and H helix regions. An equilibrium molten globule with a similar structure is formed near pH 4. Previous studies suggested that the folding intermediates are kinetically trapped states in which folding is impeded by non-native packing of the G and H helices. Fluorescence spectra of mutant proteins in which cysteine residues were introduced at several positions in the G and H helices show differential quenching of W14 fluorescence, providing direct evidence of translocation of the H helix relative to helices A and G in both the kinetic and equilibrium intermediates. Förster resonance energy transfer measurements show that a 5-({2-[(acetyl)amino]ethyl}amino)naphthalene-1-sulfonic acid acceptor coupled to K140C (helix H) is closer to Trp14 (helix A) in the equilibrium molten globule than in the native state, by a distance that is consistent with sliding of the H helix in an N-terminal direction by approximately one helical turn. Formation of an S108C–L135C disulfide prevents H helix translocation in the equilibrium molten globule by locking the G and H helices into their native register. By enforcing nativelike packing of the A, G, and H helices, the disulfide resolves local energetic frustration and facilitates transient docking of the E helix region onto the hydrophobic core but has only a small effect on the refolding rate. The apomyoglobin folding landscape is highly rugged, with several energetic bottlenecks that frustrate folding; relief of any one of the major identified bottlenecks is insufficient to speed progression to the transition state.
Co-reporter:Phillip C. Aoto, R. Bryn Fenwick, Gerard J.A. Kroon, Peter E. Wright
Journal of Magnetic Resonance 2014 246() pp: 31-35
Publication Date(Web):1 September 2014
DOI:10.1016/j.jmr.2014.06.020
•Optimal Poisson-gap sampling schemes are demonstrated to be generated infrequently.•A scoring function and software are presented to easily rank sampling schemes.•Optimal schemes lead to optimal peak intensity reconstruction.Non-uniform sampling (NUS) in NMR spectroscopy is a recognized and powerful tool to minimize acquisition time. Recent advances in reconstruction methodologies are paving the way for the use of NUS in quantitative applications, where accurate measurement of peak intensities is crucial. The presence or absence of NUS artifacts in reconstructed spectra ultimately determines the success of NUS in quantitative NMR. The quality of reconstructed spectra from NUS acquired data is dependent upon the quality of the sampling scheme. Here we demonstrate that the best performing sampling schemes make up a very small percentage of the total randomly generated schemes. A scoring method is found to accurately predict the quantitative similarity between reconstructed NUS spectra and those of fully sampled spectra. We present an easy-to-use protocol to batch generate and rank optimal Poisson-gap NUS schedules for use with 2D NMR with minimized noise and accurate signal reproduction, without the need for the creation of synthetic spectra.Download high-res image (293KB)Download full-size image
Co-reporter:James S. Fraser;R. Bryn Fenwick;Henry van den Bedem
PNAS 2014 Volume 111 (Issue 4 ) pp:E445-E454
Publication Date(Web):2014-01-28
DOI:10.1073/pnas.1323440111
Detailed descriptions of atomic coordinates and motions are required for an understanding of protein dynamics and their relation
to molecular recognition, catalytic function, and allostery. Historically, NMR relaxation measurements have played a dominant
role in the determination of the amplitudes and timescales (picosecond–nanosecond) of bond vector fluctuations, whereas high-resolution
X-ray diffraction experiments can reveal the presence of and provide atomic coordinates for multiple, weakly populated substates
in the protein conformational ensemble. Here we report a hybrid NMR and X-ray crystallography analysis that provides a more
complete dynamic picture and a more quantitative description of the timescale and amplitude of fluctuations in atomic coordinates
than is obtainable from the individual methods alone. Order parameters (S2) were calculated from single-conformer and multiconformer models fitted to room temperature and cryogenic X-ray diffraction
data for dihydrofolate reductase. Backbone and side-chain order parameters derived from NMR relaxation experiments are in
excellent agreement with those calculated from the room-temperature single-conformer and multiconformer models, showing that
the picosecond timescale motions observed in solution occur also in the crystalline state. These motions are quenched in the
crystal at cryogenic temperatures. The combination of NMR and X-ray crystallography in iterative refinement promises to provide
an atomic resolution description of the alternate conformational substates that are sampled through picosecond to nanosecond
timescale fluctuations of the protein structure. The method also provides insights into the structural heterogeneity of nonmethyl
side chains, aromatic residues, and ligands, which are less commonly analyzed by NMR relaxation measurements.
Co-reporter:Lisa M. Tuttle, H. Jane Dyson, and Peter E. Wright
Biochemistry 2013 Volume 52(Issue 20) pp:
Publication Date(Web):April 24, 2013
DOI:10.1021/bi400322e
Escherichia coli dihydrofolate reductase (DHFR) provides a paradigm for the integrated study of the role of protein dynamics in enzyme function. Previous studies of backbone and side chain dynamics have yielded unprecedented insights into the mechanism by which DHFR progresses through the structural changes that occur during its catalytic cycle. Here we report a comprehensive study of the χ1 rotamer populations of the aromatic and γ-methyl containing residues for complexes of the catalytic cycle, based on NMR measurement of 3JCγCO and 3JCγN coupling constants. We report conformational and dynamic information for eight distinct complexes, where transitions between rotamer wells may occur on a broad picosecond to millisecond time scale. This large volume of 3J data has allowed us to fit new Karplus parameterizations for aromatic side chains and to select the best available of previously determined parameters for Ile, Thr, and Val. The 3JCγCO and 3JCγN coupling constants are found to be extremely sensitive measures of side chain χ1 rotamers and to give important insights into the extent of conformational averaging. For a subset of residues in DHFR, the extent of rotamer averaging is invariant to the nature of the bound ligand, while for other residues the rotamer averaging differs in one or more complexes of the enzymatic cycle. These variable-averaging residues are generally located near the active site, but the phenomenon extends into the adenosine binding domain. For several residues, the rotamer populations in different DHFR complexes appear to depend on whether the complex is in the closed or occluded state, and some residues are exquisitely sensitive to small changes in the nature of the bound ligand.
Co-reporter:David D. Boehr, Jason R. Schnell, Dan McElheny, Sung-Hun Bae, Brendan M. Duggan, Stephen J. Benkovic, H. Jane Dyson, and Peter E. Wright
Biochemistry 2013 Volume 52(Issue 27) pp:
Publication Date(Web):June 11, 2013
DOI:10.1021/bi400563c
Correlated networks of amino acids have been proposed to play a fundamental role in allostery and enzyme catalysis. These networks of amino acids can be traced from surface-exposed residues all the way into the active site, and disruption of these networks can decrease enzyme activity. Substitution of the distal Gly121 residue in Escherichia coli dihydrofolate reductase results in an up to 200-fold decrease in the hydride transfer rate despite the fact that the residue is located 15 Å from the active-site center. In this study, nuclear magnetic resonance relaxation experiments are used to demonstrate that dynamics on the picosecond to nanosecond and microsecond to millisecond time scales are changed significantly in the G121V mutant of dihydrofolate reductase. In particular, picosecond to nanosecond time scale dynamics are decreased in the FG loop (containing the mutated residue at position 121) and the neighboring active-site loop (the Met20 loop) in the mutant compared to those of the wild-type enzyme, suggesting that these loops are dynamically coupled. Changes in methyl order parameters reveal a pathway by which dynamic perturbations can be propagated more than 25 Å across the protein from the site of mutation. All of the enzyme complexes, including the model Michaelis complex with folate and nicotinamide adenine dinucleotide phosphate bound, assume an occluded ground-state conformation, and we do not observe sampling of a higher-energy closed conformation by 15N R2 relaxation dispersion experiments. This is highly significant, because it is only in the closed conformation that the cofactor and substrate reactive centers are positioned for reaction. The mutation also impairs microsecond to millisecond time scale fluctuations that have been implicated in the release of product from the wild-type enzyme. Our results are consistent with an important role for Gly121 in controlling protein dynamics critical for enzyme function and further validate the dynamic energy landscape hypothesis of enzyme catalysis.
Co-reporter:Munehito Arai ; Josephine C. Ferreon
Journal of the American Chemical Society 2012 Volume 134(Issue 8) pp:3792-3803
Publication Date(Web):January 24, 2012
DOI:10.1021/ja209936u
Determination of affinities and binding sites involved in protein–ligand interactions is essential for understanding molecular mechanisms in biological systems. Here we combine singular value decomposition and global analysis of NMR chemical shift perturbations caused by protein–protein interactions to determine the number and location of binding sites on the protein surface and to measure the binding affinities. Using this method we show that the isolated AD1 and AD2 binding motifs, derived from the intrinsically disordered N-terminal transactivation domain of the tumor suppressor p53, both interact with the TAZ2 domain of the transcriptional coactivator CBP at two binding sites. Simulations of titration curves and line shapes show that a primary dissociation constant as small as 1–10 nM can be accurately estimated by NMR titration methods, provided that the primary and secondary binding processes are coupled. Unexpectedly, the site of binding of AD2 on the hydrophobic surface of TAZ2 overlaps with the binding site for AD1, but AD2 binds TAZ2 more tightly. The results highlight the complexity of interactions between intrinsically disordered proteins and their targets. Furthermore, the association rate of AD2 to TAZ2 is estimated to be 1.7 × 1010 M–1 s–1, approaching the diffusion-controlled limit and indicating that intrinsic disorder plus complementary electrostatics can significantly accelerate protein binding interactions.
Co-reporter:Bethany A. Buck-Koehntop;Robyn L. Stanfield;Maria A. Martinez-Yamout;Damian C. Ekiert;H. Jane Dyson;Ian A. Wilson
PNAS 2012 Volume 109 (Issue 38 ) pp:15229-15234
Publication Date(Web):2012-09-18
DOI:10.1073/pnas.1213726109
Methylation of CpG dinucleotides in DNA is a common epigenetic modification in eukaryotes that plays a central role in maintenance
of genome stability, gene silencing, genomic imprinting, development, and disease. Kaiso, a bifunctional Cys2His2 zinc finger protein implicated in tumor-cell proliferation, binds to both methylated CpG (mCpG) sites and a specific nonmethylated
DNA motif (TCCTGCNA) and represses transcription by recruiting chromatin remodeling corepression machinery to target genes.
Here we report structures of the Kaiso zinc finger DNA-binding domain in complex with its nonmethylated, sequence-specific
DNA target (KBS) and with a symmetrically methylated DNA sequence derived from the promoter region of E-cadherin. Recognition of specific bases in the major groove of the core KBS and mCpG sites is accomplished through both classical
and methyl CH···O hydrogen-bonding interactions with residues in the first two zinc fingers, whereas residues in the C-terminal
extension following the third zinc finger bind in the opposing minor groove and are required for high-affinity binding. The
C-terminal region is disordered in the free protein and adopts an ordered structure upon binding to DNA. The structures of
these Kaiso complexes provide insights into the mechanism by which a zinc finger protein can recognize mCpG sites as well
as a specific, nonmethylated regulatory DNA sequence.
Co-reporter:Gira Bhabha;Jeeyeon Lee;Damian C. Ekiert;Jongsik Gam;Ian A. Wilson;H. Jane Dyson;Stephen J. Benkovic
Science 2011 Volume 332(Issue 6026) pp:234-238
Publication Date(Web):08 Apr 2011
DOI:10.1126/science.1198542
An Escherichia coli dihydrofolate reductase mutant is catalytically defective, because motions in the active site are impaired.
Co-reporter:Derrick W. Meinhold
PNAS 2011 Volume 108 (Issue 22 ) pp:9078-9083
Publication Date(Web):2011-05-31
DOI:10.1073/pnas.1105682108
Detailed understanding of protein function and malfunction hinges on the ability to characterize transiently populated states
and the transitions between them. Here, we use 15N, , and 13CO NMR R2 relaxation dispersion to investigate spontaneous unfolding and refolding events of native apomyoglobin. Above pH 5.0, dispersion
is dominated by processes involving fluctuations of the F-helix region, which is invisible in NMR spectra. Measurements of
R2 dispersion for residues contacted by the F-helix region in the native (N) structure reveal a transient state formed by local
unfolding of helix F and undocking from the protein core. A similar state was detected at pH 4.75–4.95 and determined to be
an on-pathway intermediate (I1) in a linear three-state unfolding scheme (N⇆I1⇆MG) leading to a transiently populated molten
globule (MG) state. The slowest steps in unfolding and refolding are N → I1 (36 s-1) and MG → I1 (26 s-1), respectively. Differences in chemical shift between N and I1 are very small, except in regions adjacent to helix F, showing
that their core structures are similar. Chemical shift changes between the N and MG states, obtained from R2 dispersion, reveal that the transient MG state is structurally similar to the equilibrium MG observed previously at high
temperature and low pH. Analysis of MG state chemical shifts shows the location of residual helical structure in the transient
intermediate and identifies regions that unfold or rearrange into nonnative structure during the N → MG transition. The experiments
also identify regions of energetic frustration that “crack” during unfolding and impede the refolding process.
Co-reporter:Chul Won Lee, Maria A. Martinez-Yamout, H. Jane Dyson, and Peter E. Wright
Biochemistry 2010 Volume 49(Issue 46) pp:
Publication Date(Web):October 20, 2010
DOI:10.1021/bi1012996
The activity and stability of the tumor suppressor p53 are regulated by interactions with key cellular proteins such as MDM2 and CBP/p300. The transactivation domain (TAD) of p53 contains two subdomains (AD1 and AD2) and interacts directly with the N-terminal domain of MDM2 and with several domains of CBP/p300. Here we report the NMR structure of the full-length p53 TAD in complex with the nuclear coactivator binding domain (NCBD) of CBP. Both the p53 TAD and NCBD are intrinsically disordered and fold synergistically upon binding, as evidenced by the observed increase in helicity and increased level of dispersion of the amide proton resonances. The p53 TAD folds to form a pair of helices (denoted Pα1 and Pα2), which extend from Phe19 to Leu25 and from Pro47 to Trp53, respectively. In the complex, the NCBD forms a bundle of three helices (Cα1, residues 2066−2075; Cα2, residues 2081−2092; and Cα3, residues 2095−2105) with a hydrophobic groove into which p53 helices Pα1 and Pα2 dock. The polypeptide chain between the p53 helices remains flexible and makes no detectable intermolecular contacts with the NCBD. Complex formation is driven largely by hydrophobic contacts that form a stable intermolecular hydrophobic core. A salt bridge between D49 of p53 and R2105 of NCBD may contribute to the binding specificity. The structure provides the first insights into simultaneous binding of the AD1 and AD2 motifs to a target protein.
Co-reporter:Chul Won Lee;Josephine C. Ferreon;Allan Chris M. Ferreon;Munehito Arai
PNAS 2010 107 (45 ) pp:19290-19295
Publication Date(Web):2010-11-09
DOI:10.1073/pnas.1013078107
The transcriptional activity of p53 is regulated by a cascade of posttranslational modifications. Although acetylation of
p53 by CREB-binding protein (CBP)/p300 is known to be indispensable for p53 activation, the role of phosphorylation, and in
particular multisite phosphorylation, in activation of CBP/p300-dependent p53 transcriptional pathways remains unclear. We
investigated the role of single site and multiple site phosphorylation of the p53 transactivation domain in mediating its
interaction with CBP and with the ubiquitin ligase HDM2. Phosphorylation at Thr18 functions as an on/off switch to regulate
binding to the N-terminal domain of HDM2. In contrast, binding to CBP is modulated by the extent of p53 phosphorylation; addition
of successive phosphoryl groups enhances the affinity for the TAZ1, TAZ2, and KIX domains of CBP in an additive manner. Activation
of p53-dependent transcriptional pathways requires that p53 compete with numerous cellular transcription factors for binding
to limiting amounts of CBP/p300. Multisite phosphorylation represents a mechanism for a graded p53 response, with each successive
phosphorylation event resulting in increasingly efficient recruitment of CBP/p300 to p53-regulated transcriptional programs,
in the face of competition from cellular transcription factors. Multisite phosphorylation thus acts as a rheostat to enhance
binding to CBP/p300 and provides a plausible mechanistic explanation for the gradually increasing p53 response observed following
prolonged or severe genotoxic stress.
Co-reporter:Dan McElheny;David D. Boehr;H. Jane Dyson
PNAS 2010 Volume 107 (Issue 4 ) pp:1373-1378
Publication Date(Web):2010-01-26
DOI:10.1073/pnas.0914163107
Enzyme catalysis can be described as progress over a multi-dimensional energy landscape where ensembles of interconverting
conformational substates channel the enzyme through its catalytic cycle. We applied NMR relaxation dispersion to investigate
the role of bound ligands in modulating the dynamics and energy landscape of Escherichia coli dihydrofolate reductase to obtain insights into the mechanism by which the enzyme efficiently samples functional conformations
as it traverses its reaction pathway. Although the structural differences between the occluded substrate binary complexes
and product ternary complexes are very small, there are substantial differences in protein dynamics. Backbone fluctuations
on the μs-ms timescale in the cofactor binding cleft are similar for the substrate and product binary complexes, but fluctuations
on this timescale in the active site loops are observed only for complexes with substrate or substrate analog and are not
observed for the binary product complex. The dynamics in the substrate and product binary complexes are governed by quite
different kinetic and thermodynamic parameters. Analogous dynamic differences in the E:THF:NADPH and E:THF:NADP+ product ternary complexes are difficult to rationalize from ground-state structures. For both of these complexes, the nicotinamide
ring resides outside the active site pocket in the ground state. However, they differ in the structure, energetics, and dynamics
of accessible higher energy substates where the nicotinamide ring transiently occupies the active site. Overall, our results
suggest that dynamics in dihydrofolate reductase are exquisitely “tuned” for every intermediate in the catalytic cycle; structural
fluctuations efficiently channel the enzyme through functionally relevant conformational space.
Co-reporter:Sung-Hun Bae ; H. Jane Dyson
Journal of the American Chemical Society 2009 Volume 131(Issue 19) pp:6814-6821
Publication Date(Web):April 24, 2009
DOI:10.1021/ja809687r
For well-structured, rigid proteins, the prediction of rotational tumbling time (τc) using atomic coordinates is reasonably accurate, but is inaccurate for proteins with long unstructured sequences. Under physiological conditions, many proteins contain long disordered segments that play important regulatory roles in fundamental biological events including signal transduction and molecular recognition. Here we describe an ensemble approach to the boundary element method that accurately predicts τc for such proteins by introducing two layers of molecular surfaces whose correlated velocities decay exponentially with distance. Reliable prediction of τc will help to detect intra- and intermolecular interactions and conformational switches between more ordered and less ordered states of the disordered segments. The method has been extensively validated using 12 reference proteins with 14 to 103 disordered residues at the N- and/or C-terminus and has been successfully employed to explain a set of published results on a system that incorporates a conformational switch.
Co-reporter:Chul Won Lee, Munehito Arai, Maria A. Martinez-Yamout, H. Jane Dyson and Peter E. Wright
Biochemistry 2009 Volume 48(Issue 10) pp:
Publication Date(Web):February 16, 2009
DOI:10.1021/bi802055v
Molecular interactions between the tumor suppressor p53 and the transcriptional coactivators CBP/p300 are critical for the regulation of p53 transactivation and stability. The transactivation domain (TAD) of p53 binds directly to several CBP/p300 domains (TAZ1, TAZ2, NCBD, and KIX). Here we map the interaction between the p53 TAD and the CBP KIX domain using isothermal titration calorimetry and NMR spectroscopy. KIX is a structural domain in CBP/p300 that can simultaneously bind two polypeptide ligands, such as the activation domain of MLL and the kinase-inducible activation domain (pKID) of CREB, using distinct interaction surfaces. The p53 TAD consists of two subdomains (AD1 and AD2); peptides corresponding to the isolated AD1 and AD2 subdomains interact with KIX with relatively low affinity, but a longer peptide containing both subdomains binds KIX tightly. In the context of the full-length p53 TAD, AD1 and AD2 bind synergistically to KIX. Mapping of the chemical shift perturbations onto the structure of KIX shows that isolated AD1 and AD2 peptides bind to both the MLL and pKID sites. Spin-labeling experiments show that the complex of the full-length p53 TAD with KIX is disordered, with the AD1 and AD2 subdomains each interacting with both the MLL and pKID binding surfaces. Phosphorylation of the p53 TAD at Thr18 or Ser20 increases the KIX binding affinity. The affinity is further enhanced by simultaneous phosphorylation of Thr18 and Ser20, and the specificity of the interaction is increased. The p53 TAD simultaneously occupies the two distinct sites that have been identified on the CBP KIX domain and efficiently competes for these sites with other known KIX-binding transcription factors.
Co-reporter:Josephine C. Ferreon;Maria A. Martinez-Yamout;H. Jane Dyson
PNAS 2009 Volume 106 (Issue 32 ) pp:13260-13265
Publication Date(Web):2009-08-11
DOI:10.1073/pnas.0906770106
The adenovirus early region 1A (E1A) oncoprotein mediates cell transformation by deregulating host cellular processes and
activating viral gene expression by recruitment of cellular proteins that include cyclic-AMP response element binding (CREB)
binding protein (CBP)/p300 and the retinoblastoma protein (pRb). While E1A is capable of independent interaction with CBP/p300
or pRb, simultaneous binding of both proteins is required for maximal biological activity. To obtain insights into the mechanism
by which E1A hijacks the cellular transcription machinery by competing with essential transcription factors for binding to
CBP/p300, we have determined the structure of the complex between the transcriptional adaptor zinc finger-2 (TAZ2) domain
of CBP and the conserved region-1 (CR1) domain of E1A. The E1A CR1 domain is unstructured in the free state and upon binding
folds into a local helical structure mediated by an extensive network of intermolecular hydrophobic contacts. By NMR titrations,
we show that E1A efficiently competes with the N-terminal transactivation domain of p53 for binding to TAZ2 and that pRb interacts
with E1A at 2 independent sites located in CR1 and CR2. We show that pRb and the CBP TAZ2 domain can bind simultaneously to
the CR1 site of E1A to form a ternary complex and propose a structural model for the pRb:E1A:CBP complex on the basis of published
x-ray data for homologous binary complexes. These observations reveal the molecular basis by which E1A inhibits p53-mediated
transcriptional activation and provide a rationale for the efficiency of cellular transformation by the adenoviral E1A oncoprotein.
Co-reporter:Josephine C. Ferreon;Chul Won Lee;Munehito Arai;Maria A. Martinez-Yamout;H. Jane Dyson
PNAS 2009 Volume 106 (Issue 16 ) pp:6591-6596
Publication Date(Web):2009-04-21
DOI:10.1073/pnas.0811023106
The tumor suppressor activity of p53 is regulated by interactions with the ubiquitin ligase HDM2 and the general transcriptional
coactivators CBP and p300. Using NMR spectroscopy and isothermal titration calorimetry, we have dissected the binding interactions
between the N-terminal transactivation domain (TAD) of p53, the TAZ1, TAZ2, KIX, and nuclear receptor coactivator binding
domains of CBP, and the p53-binding domain of HDM2. The p53 TAD contains amphipathic binding motifs within the AD1 and AD2
regions that mediate interactions with CBP and HDM2. Binding of the p53 TAD to CBP domains is dominated by interactions with
AD2, although the affinity is enhanced by additional interactions with AD1. In contrast, binding of p53 TAD to HDM2 is mediated
primarily by AD1. The p53 TAD can bind simultaneously to HDM2 (through AD1) and to any one of the CBP domains (through AD2)
to form a ternary complex. Phosphorylation of p53 at T18 impairs binding to HDM2 and enhances affinity for the CBP KIX domain.
Multisite phosphorylation of the p53 TAD at S15, T18, and S20 leads to increased affinity for the TAZ1 and KIX domains of
CBP. These observations suggest a mechanism whereby HDM2 and CBP/p300 function synergistically to regulate the p53 response.
In unstressed cells, CBP/p300, HDM2 and p53 form a ternary complex that promotes polyubiquitination and degradation of p53.
After cellular stress and DNA damage, p53 becomes phosphorylated at T18 and other residues in the AD1 region, releases HDM2
and binds preferentially to CBP/p300, leading to stabilization and activation of p53.
Co-reporter:David D. Boehr, H. Jane Dyson and Peter E. Wright
Biochemistry 2008 Volume 47(Issue 35) pp:
Publication Date(Web):August 9, 2008
DOI:10.1021/bi801102e
The catalytic cycle of an enzyme is frequently associated with conformational changes that may limit maximum catalytic throughput. In Escherichia coli dihydrofolate reductase, release of the tetrahydrofolate (THF) product is the rate-determining step under physiological conditions and is associated with an “occluded” to “closed” conformational change. In this study, we demonstrate that in dihydrofolate reductase the closed to occluded conformational change in the product ternary complex (E·THF·NADP+) also gates progression through the catalytic cycle. Using NMR relaxation dispersion, we have measured the temperature and pH dependence of microsecond to millisecond time scale backbone dynamics of the occluded E·THF·NADP+ complex. Our studies indicate the presence of three independent dynamic regions, associated with the active-site loops, the cofactor binding cleft, and the C-terminus and an adjacent loop, which fluctuate into discrete conformational substates with different kinetic and thermodynamic parameters. The dynamics of the C-terminally associated region is pH-dependent (pKa < 6), but the dynamics of the active-site loops and cofactor binding cleft are pH-independent. The active-site loop dynamics access a closed conformation, and the accompanying closed to occluded rate constant is comparable to the maximum pH-independent hydride transfer rate constant. Together, these results strongly suggest that the closed to occluded conformational transition in the product ternary complex is a prerequisite for progression through the catalytic cycle and that the rate of this process places an effective limit on the maximum rate of the hydride transfer step.
Co-reporter:Chiaki Nishimura;Takanori Uzawa;Koichiro Ishimori;Shuji Akiyama;Satoshi Takahashi;H. Jane Dyson
PNAS 2008 Volume 105 (Issue 37 ) pp:13859-13864
Publication Date(Web):2008-09-16
DOI:10.1073/pnas.0804033105
The earliest steps in the folding of proteins are complete on an extremely rapid time scale that is difficult to access experimentally.
We have used rapid-mixing quench-flow methods to extend the time resolution of folding studies on apomyoglobin and elucidate
the structural and dynamic features of members of the ensemble of intermediate states that are populated on a submillisecond
time scale during this process. The picture that emerges is of a continuum of rapidly interconverting states. Even after only
0.4 ms of refolding time a compact state is formed that contains major parts of the A, G, and H helices, which are sufficiently
well folded to protect amides from exchange. The B, C, and E helix regions fold more slowly and fluctuate rapidly between
open and closed states as they search docking sites on this core; the secondary structure in these regions becomes stabilized
as the refolding time is increased from 0.4 to 6 ms. No further stabilization occurs in the A, G, H core at 6 ms of folding
time. These studies begin to time-resolve a progression of compact states between the fully unfolded and native folded states
and confirm the presence an ensemble of intermediates that interconvert in a hierarchical sequence as the protein searches
conformational space on its folding trajectory.
Co-reporter:Michael A. Lietzow;H. Jane Dyson;Daniel J. Felitsky
PNAS 2008 Volume 105 (Issue 17 ) pp:6278-6283
Publication Date(Web):2008-04-29
DOI:10.1073/pnas.0710641105
The primary driving force for protein folding is the sequestration of hydrophobic side chains from solvent water, but the
means whereby the amino acid sequence directs the folding process to form the correct final folded state is not well understood.
Measurements of NMR line broadening in spin-labeled samples of unfolded apomyoglobin at pH 2.3 have been used to derive a
quantitative model for transient hydrophobic interactions between various sites in the polypeptide chain, as would occur during
the initiation of protein folding. Local clusters of residues with high values for the parameter “average area buried upon
folding” (AABUF) form foci not only for local contacts but for long-range interactions, the relative frequencies of which
can be understood in terms of differences in the extent of reduction in chain configurational entropy that occurs upon formation
of nonlocal contacts. These results complement the striking correlation previously observed between the kinetic folding process
of apomyoglobin and the AABUF of its amino acid sequence [Nishimura C, Lietzow MA, Dyson HJ, Wright PE (2005) J Mol Biol 351:383–392]. For the acid-unfolded states of apomyoglobin, our approach identifies multiple distinct hydrophobic clusters
of differing thermodynamic stability. The most structured of these clusters, although sparsely populated, have both native-like
and nonnative character; the specificity of the transient long-range contacts observed in these states suggests that they
play a key role in initiating chain collapse and folding.
Co-reporter:Kenji Sugase,
H. Jane Dyson
&
Peter E. Wright
Nature 2007 447(7147) pp:1021
Publication Date(Web):2007-05-23
DOI:10.1038/nature05858
Protein folding and binding are analogous processes, in which the protein ‘searches’ for favourable intramolecular or intermolecular interactions on a funnelled energy landscape1, 2. Many eukaryotic proteins are disordered under physiological conditions, and fold into ordered structures only on binding to their cellular targets3, 4, 5, 6. The mechanism by which folding is coupled to binding is poorly understood, but it has been hypothesized on theoretical grounds that the binding kinetics may be enhanced by a ‘fly-casting’ effect, where the disordered protein binds weakly and non-specifically to its target and folds as it approaches the cognate binding site7. Here we show, using NMR titrations and 15N relaxation dispersion, that the phosphorylated kinase inducible activation domain (pKID) of the transcription factor CREB forms an ensemble of transient encounter complexes on binding to the KIX domain of the CREB binding protein. The encounter complexes are stabilized primarily by non-specific hydrophobic contacts, and evolve by way of an intermediate to the fully bound state without dissociation from KIX. The carboxy-terminal helix of pKID is only partially folded in the intermediate, and becomes stabilized by intermolecular interactions formed in the final bound state. Future applications of our method will provide new understanding of the molecular mechanisms by which intrinsically disordered proteins perform their diverse biological functions.
Co-reporter:David D. Boehr;Dan McElheny;H. Jane Dyson
Science 2006 Vol 313(5793) pp:1638-1642
Publication Date(Web):15 Sep 2006
DOI:10.1126/science.1130258
Abstract
We used nuclear magnetic resonance relaxation dispersion to characterize higher energy conformational substates of Escherichia coli dihydrofolate reductase. Each intermediate in the catalytic cycle samples low-lying excited states whose conformations resemble the ground-state structures of preceding and following intermediates. Substrate and cofactor exchange occurs through these excited substates. The maximum hydride transfer and steady-state turnover rates are governed by the dynamics of transitions between ground and excited states of the intermediates. Thus, the modulation of the energy landscape by the bound ligands funnels the enzyme through its reaction cycle along a preferred kinetic path.
Co-reporter:Dan McElheny;Jason R. Schnell;Jonathan C. Lansing;H. Jane Dyson;
Proceedings of the National Academy of Sciences 2005 102(14) pp:5032-5037
Publication Date(Web):March 28, 2005
DOI:10.1073/pnas.0500699102
Dynamic processes are implicit in the catalytic function of all enzymes. To obtain insights into the relationship between
the dynamics and thermodynamics of protein fluctuations and catalysis, we have measured millisecond time scale motions in
the enzyme dihydrofolate reductase using NMR relaxation methods. Studies of a ternary complex formed from the substrate analog
folate and oxidized NADP+ cofactor revealed conformational exchange between a ground state, in which the active site loops adopt a closed conformation,
and a weakly populated (4.2% at 30°C) excited state with the loops in the occluded conformation. Fluctuations between these
states, which involve motions of the nicotinamide ring of the cofactor into and out of the active site, occur on a time scale
that is directly relevant to the structural transitions involved in progression through the catalytic cycle.
Co-reporter:Chiaki Nishimura;H. Jane Dyson
PNAS 2005 102 (13 ) pp:4765-4770
Publication Date(Web):2005-03-29
DOI:10.1073/pnas.0409538102
Hydrogen/deuterium exchange followed by trapping of the labeled species in the aprotic solvent DMSO has been used to elucidate
structure in both the burst-phase molten globule-folding intermediate of apomyoglobin and in an equilibrium intermediate that
models the kinetic intermediate. Precise estimates can be made of exchange times in an interrupted exchange-out experiment
at pH 4 followed by analysis in DMSO solution, giving extensive sequence-specific information about the structure of the equilibrium
intermediate. In addition, the use of DMSO as a solvent for NMR measurements after quench-flow pH-pulse labeling experiments
gives a greatly increased data set for the elucidation of the kinetic folding pathway. Interestingly, differences are observed
in some regions of apomyoglobin between the equilibrium and kinetic intermediates. These differences are quantitative rather
than qualitative; that is, the overall patterns of labeling and secondary structure formation remain similar between the two
species. However, local differences are observed, which probably reflect the difference in the solution conditions for the
equilibrium experiment (pH 4) vs. the kinetic experiment (pH 6) and the change in the status of the stabilizing hydrogen bond
between the side chains of His-24 and His-119.
Co-reporter:
Nature Structural and Molecular Biology 2004 11(3) pp:257-264
Publication Date(Web):08 February 2004
DOI:10.1038/nsmb738
The tandem zinc finger (TZF) domain of the protein TIS11d binds to the class II AU-rich element (ARE) in the 3' untranslated region (3' UTR) of target mRNAs and promotes their deadenylation and degradation. The NMR structure of the TIS11d TZF domain bound to the RNA sequence 5'-UUAUUUAUU-3' comprises a pair of novel CCCH fingers of type CX8CX5CX3H separated by an 18-residue linker. The two TIS11d zinc fingers bind in a symmetrical fashion to adjacent 5'-UAUU-3' subsites on the single-stranded RNA via a combination of electrostatic and hydrogen-bonding interactions, with intercalative stacking between conserved aromatic side chains and the RNA bases. Sequence specificity in RNA recognition is achieved by a network of intermolecular hydrogen bonds, mostly between TIS11d main-chain functional groups and the Watson-Crick edges of the bases. The TIS11d structure provides insights into the RNA-binding functions of this large family of CCCH zinc finger proteins.
Co-reporter:Stephen J. Demarest;Maria Martinez-Yamout;John Chung;Hongwu Chen;Wei Xu;H. Jane Dyson;Ronald M. Evans
Nature 2002 415(6871) pp:549-553
Publication Date(Web):2002-01-31
DOI:10.1038/415549a
Nuclear hormone receptors are ligand-activated transcription factors that regulate the expression of genes that are essential for development, reproduction and homeostasis1. The hormone response is mediated through recruitment of p160 receptor coactivators and the general transcriptional coactivator CBP/p300, which function synergistically to activate transcription2. These coactivators exhibit intrinsic histone acetyltransferase activity, function in the remodelling of chromatin, and facilitate the recruitment of RNA polymerase II and the basal transcription machinery3. The activities of the p160 coactivators are dependent on CBP. Both coactivators are essential for proper cell-cycle control, differentiation and apoptosis, and are implicated in cancer and other diseases4, 5, 6, 7. To elucidate the molecular basis of assembling the multiprotein activation complex, we undertook a structural and thermodynamic analysis of the interaction domains of CBP and the activator for thyroid hormone and retinoid receptors8. Here we show that although the isolated domains are intrinsically disordered, they combine with high affinity to form a cooperatively folded helical heterodimer. Our study uncovers a unique mechanism, called ‘synergistic folding’, through which p160 coactivators recruit CBP/p300 to allow transmission of the hormonal signal to the transcriptional machinery.
Co-reporter:Roberto N. De Guzman;Maria Martinez-Yamout;Sonja A. Dames;H. Jane Dyson
PNAS 2002 Volume 99 (Issue 8 ) pp:5271-5276
Publication Date(Web):2002-04-16
DOI:10.1073/pnas.082121399
The cellular response to low tissue oxygen concentrations is mediated by the hypoxia-inducible transcription factor HIF-1.
Under hypoxic conditions, HIF-1 activates transcription of critical adaptive genes by recruitment of the general coactivators
CBP/p300 through interactions with its α-subunit (Hif-1α). Disruption of the Hif-1α/p300 interaction has been linked to attenuation
of tumor growth. To delineate the structural basis for this interaction, we have determined the solution structure of the
complex between the carboxy-terminal activation domain (CAD) of Hif-1α and the zinc-binding TAZ1 (CH1) motif of cyclic-AMP
response element binding protein (CREB) binding protein (CBP). Despite the overall similarity of the TAZ1 structure to that
of the TAZ2 (part of the CH3) domain of CBP, differences occur in the packing of helices that can account for differences
in specificity. The unbound CAD is intrinsically disordered and remains relatively extended upon binding, wrapping almost
entirely around the TAZ1 domain in a groove through much of its surface. Three short helices are formed upon binding, stabilized
by intermolecular interactions. The Asn-803 side chain, which functions as a hypoxic switch, is located on the second of these
helices and is buried in the molecular interface. The third helix of the Hif-1α CAD docks in a deep hydrophobic groove in
TAZ1, providing extensive intermolecular hydrophobic interactions that contribute to the stability of the complex. The structure
of this complex provides new insights into the mechanism through which Hif-1α recruits CBP/p300 in response to hypoxia.
Co-reporter:John H. Laity;H. Jane Dyson
PNAS 2000 Volume 97 (Issue 22 ) pp:11932-11935
Publication Date(Web):2000-10-24
DOI:10.1073/pnas.97.22.11932
Alternate splicing, leading to the insertion of the tripeptide KTS
in the linker between the third and fourth C2H2
zinc fingers, changes both the DNA-binding function and the subnuclear
localization of the Wilms' tumor suppressor protein (WT1). We
have used NMR relaxation experiments to determine the molecular
basis for the differing DNA recognition properties of the WT1–KTS and
WT1+KTS isoforms. Our results show that the KTS insertion
increases the flexibility of the linker between fingers 3 and 4 and
abrogates binding of the fourth zinc finger to its cognate site in the
DNA major groove. This represents a mechanism whereby a single
zinc-finger gene can be used, through alternate splicing, to fulfill
different functions in the cell.
Co-reporter:
Nature Structural and Molecular Biology 2000 7(8) pp:679-686
Publication Date(Web):
DOI:10.1038/77985
To test the hypothesis that the folding pathways of evolutionarily related proteins with similar three-dimensional structures but widely different sequences should be similar, the folding pathway of apoleghemoglobin has been characterized using stopped-flow circular dichroism, heteronuclear NMR pulse labeling techniques and mass spectrometry. The pathway of folding was found to differ significantly from that of a protein of the same family, apomyoglobin, although both proteins appear to fold through helical burst phase intermediates. For leghemoglobin, the burst phase intermediate exhibits stable helical structure in the G and H helices, together with a small region in the center of the E helix. The A and B helices are not stabilized until later stages of the folding process. The structure of the burst phase folding intermediate thus differs from that of apomyoglobin, in which stable helical structure is formed in the A, B, G and H helix regions.
Co-reporter:
Nature Structural and Molecular Biology 1998 5(7s) pp:499 - 503
Publication Date(Web):
DOI:10.1038/739
Co-reporter:Peter E. Wright
Journal of Molecular Biology (9 January 2009) Volume 385(Issue 1) pp:1-2
Publication Date(Web):9 January 2009
DOI:10.1016/j.jmb.2008.12.013
Co-reporter:Raphael Stoll, Brian M. Lee, Erik W. Debler, John H. Laity, ... Peter E. Wright
Journal of Molecular Biology (5 October 2007) Volume 372(Issue 5) pp:1227-1245
Publication Date(Web):5 October 2007
DOI:10.1016/j.jmb.2007.07.017
The zinc finger domain of the Wilms tumor suppressor protein (WT1) contains four canonical Cys2His2 zinc fingers. WT1 binds preferentially to DNA sequences that are closely related to the EGR-1 consensus site. We report the structure determination by both X-ray crystallography and NMR spectroscopy of the WT1 zinc finger domain in complex with DNA. The X-ray structure was determined for the complex with a cognate 14 base-pair oligonucleotide, and composite X-ray/NMR structures were determined for complexes with both the 14 base-pair and an extended 17 base-pair DNA. This combined approach allowed unambiguous determination of the position of the first zinc finger, which is influenced by lattice contacts in the crystal structure. The crystal structure shows the second, third and fourth zinc finger domains inserted deep into the major groove of the DNA where they make base-specific interactions. The DNA duplex is distorted in the vicinity of the first zinc finger, with a cytidine twisted and tilted out of the base stack to pack against finger 1 and the tip of finger 2. By contrast, the composite X-ray/NMR structures show that finger 1 continues to follow the major groove in the solution complexes. However, the orientation of the helix is non-canonical, and the fingertip and the N terminus of the helix project out of the major groove; as a consequence, the zinc finger side-chains that are commonly involved in base recognition make no contact with the DNA. We conclude that finger 1 helps to anchor WT1 to the DNA by amplifying the binding affinity although it does not contribute significantly to binding specificity. The structures provide molecular level insights into the potential consequences of mutations in zinc fingers 2 and 3 that are associated with Denys-Drash syndrome and nephritic syndrome. The mutations are of two types, and either destabilize the zinc finger structure or replace key base contact residues.
Co-reporter:Chiaki Nishimura, H. Jane Dyson, Peter E. Wright
Journal of Molecular Biology (5 August 2011) Volume 411(Issue 1) pp:248-263
Publication Date(Web):5 August 2011
DOI:10.1016/j.jmb.2011.05.028
The F helix region of sperm whale apomyoglobin is disordered, undergoing conformational fluctuations between a folded helical conformation and one or more locally unfolded states. To examine the effects of F helix stabilization on the folding pathway of apomyoglobin, we have introduced mutations to augment intrinsic helical structure in the F helix of the kinetic folding intermediate and to increase its propensity to fold early in the pathway, using predictions based on plots of the average area buried upon folding (AABUF) derived from the primary sequence. Two mutant proteins were prepared: a double mutant, P88K/S92K (F2), and a quadruple mutant, P88K/A90L/S92K/A94L (F4). Whereas the AABUF for F2 predicts that the F helix will not fold early in the pathway, the F helix in F4 shows a significantly increased AABUF and is therefore predicted to fold early. Protection of amide protons by formation of hydrogen-bonded helical structure during the early folding events has been analyzed by pH-pulse labeling. Consistent with the AABUF prediction, many of the F helix residues for F4 are significantly protected in the kinetic intermediate but are not protected in the F2 mutant. F4 folds via a kinetically trapped burst-phase intermediate that contains stabilized secondary structure in the A, B, F, G, and H helix regions. Rapid folding of the F helix stabilizes the central core of the misfolded intermediate and inhibits translocation of the H helix back to its native position, thereby decreasing the overall folding rate.Graphical AbstractDownload high-res image (182KB)Download full-size imageResearch Highlights► Mutant apomyoglobins were designed to stabilize the F helix. ► F2 (P88K/S92K) and F4 (P88K/A90L/S92K/A94L) stabilized the F helix at neutral pH. ► F4 also stabilized the F helix in the kinetic and equilibrium folding intermediates. ► F4 folds more slowly due to stability of nonnative structure in the intermediate. ► Results are consistent with the AABUF (hydrophobic burial) hypothesis.
Co-reporter:Brian M. Lee, Bethany A. Buck-Koehntop, Maria A. Martinez-Yamout, H. Jane Dyson, Peter E. Wright
Journal of Molecular Biology (31 August 2007) Volume 371(Issue 5) pp:1274-1289
Publication Date(Web):31 August 2007
DOI:10.1016/j.jmb.2007.06.021
Churchill is a zinc-containing protein that is involved in neural induction during embryogenesis. At the time of its discovery, it was thought on the basis of sequence alignment to contain two zinc fingers of the C4 type. Further, binding of an N-terminal GST-Churchill fusion protein to a particular DNA sequence was demonstrated by immunoprecipitation selection assay, suggesting that Churchill may function as a transcriptional regulator by sequence-specific DNA binding. We show by NMR solution structure determination that, far from containing canonical C4 zinc fingers, the protein contains three bound zinc ions in novel coordination sites, including an unusual binuclear zinc cluster. The secondary structure of Churchill is also unusual, consisting of a highly solvent-exposed single-layer β-sheet. Hydrogen-deuterium exchange and backbone relaxation measurements reveal that Churchill is unusually dynamic on a number of time scales, with the exception of regions surrounding the zinc coordinating sites, which serve to stabilize the otherwise unstructured N terminus and the single-layer β-sheet. No binding of Churchill to the previously identified DNA sequence could be detected, and extensive searches using DNA sequence selection techniques could find no other DNA sequence that was bound by Churchill. Since the N-terminal amino acids of Churchill form part of the zinc-binding motif, the addition of a fusion protein at the N terminus causes loss of zinc and unfolding of Churchill. This observation most likely explains the published DNA-binding results, which would arise due to non-specific interaction of the unfolded protein in the immunoprecipitation selection assay. Since Churchill does not appear to bind DNA, we suggest that it may function in embryogenesis as a protein-interaction factor.
Co-reporter:Kwang Hun Lim, H. Jane Dyson, Jeffery W. Kelly, Peter E. Wright
Journal of Molecular Biology (25 March 2013) Volume 425(Issue 6) pp:977-988
Publication Date(Web):25 March 2013
DOI:10.1016/j.jmb.2013.01.008
The process of transthyretin (TTR) misfolding and aggregation, including amyloid formation, appears to cause a number of degenerative diseases. During amyloid formation, the native protein undergoes a tetramer-to-folded monomer transition, followed by local unfolding of the monomer to an assembly-competent amyloidogenic intermediate. Here we use NMR relaxation dispersion to probe conformational exchange at physiological pH between native monomeric TTR (the F87M/L110M variant) and a small population of a transiently formed amyloidogenic intermediate. The dispersion experiments show that a majority of the residues in the β-sheet containing β-strands D, A, G, and H undergo conformational fluctuations on microsecond-to-millisecond timescales. Exchange broadening is greatest for residues in the outer β-strand H, which hydrogen bonds to β-strand H′ of a neighboring subunit in the tetramer, but the associated structural fluctuations propagate across the entire β-sheet. Fluctuations in the other β-sheet are limited to the outer β-strand F, which packs against strand F′ in the tetramer, while the B, C, and E β-strands of this sheet remain stable. The structural changes were also investigated under more forcing amyloidogenic conditions (pH 6.4–3.7), where β-strand D and regions of the D–E and E–F loops were additionally destabilized, increasing the population of the amyloidogenic intermediate and accelerating amyloid formation. Strands B, C, and E appear to maintain native-like conformations in the partially unfolded, amyloidogenic state of wild-type TTR. In the case of the protective mutant T119M, the conformational fluctuations are suppressed under both physiological and mildly acidic conditions, indicating that the dynamic properties of TTR correlate well with its aggregation propensity.Download high-res image (100KB)Download full-size imageHighlights► We report relaxation dispersion measurements of monomeric TTR. ► Monomeric TTR shows microsecond-to-millisecond timescale fluctuations at neutral pH. ► The fluctuations are localized to one of the two TTR β-sheets. ► Enhanced amyloidogenic conditions at lower pH show increased fluctuation. ► These conformational fluctuations are abolished in the protective mutant T119M.
Co-reporter:Bethany A. Buck-Koehntop, Maria A. Martinez-Yamout, H. Jane Dyson, Peter E. Wright
FEBS Letters (23 March 2012) Volume 586(Issue 6) pp:734-739
Publication Date(Web):23 March 2012
DOI:10.1016/j.febslet.2012.01.045
Kaiso is a Cys2His2 zinc finger (ZF) protein that mediates methyl-CpG-dependent and sequence-specific transcriptional repression. As a first step towards elucidating the structural and molecular basis for recognition of these disparate DNA sequences, the minimal binding region of Kaiso was identified and optimal DNA sequences for high-affinity interactions were characterized. Contrary to previous findings, Kaiso requires all three zinc fingers plus adjacent protein regions for DNA recognition. An N-terminal extension contributes to structural stability, while an extended C-terminal region augments DNA binding. Complexes formed between the optimized Kaiso construct and both DNA sequences are suitable for future structural evaluation.Highlights► We have identified minimal Kaiso constructs for high-affinity DNA binding. ► All three Kaiso zinc fingers are required for binding. ► Extension of the sequence at the N-terminus increases stability of the protein. ► Extension of the sequence at the C-terminus augments DNA affinity.
Co-reporter:Chiaki Nishimura, H. Jane Dyson, Peter E. Wright
Journal of Molecular Biology (2 May 2008) Volume 378(Issue 3) pp:715-725
Publication Date(Web):2 May 2008
DOI:10.1016/j.jmb.2008.03.025
An important question in protein folding is whether molten globule states formed under equilibrium conditions are good structural models for kinetic folding intermediates. The structures of the kinetic and equilibrium intermediates in the folding of the plant globin apoleghemoglobin have been compared at high resolution by quench-flow pH-pulse labeling and interrupted hydrogen/deuterium exchange analyzed in dimethyl sulfoxide. Unlike its well studied homolog apomyoglobin, where the equilibrium and kinetic intermediates are quite similar, there are striking structural differences between the intermediates formed by apoleghemoglobin. In the kinetic intermediate, formed during the burst phase of the quench-flow experiment, protected amides and helical structure are found mainly in the regions corresponding to the G and H helices of the folded protein, and in parts of the E helix and CE loop regions, whereas in the equilibrium intermediate, amide protection and helical structure are seen in parts of the A and B helix regions, as well as in the G and H regions, and the E helix remains largely unfolded. These results suggest that the structure of the molten globule intermediate of apoleghemoglobin is more plastic than that of apomyoglobin, so that it is readily transformed depending on the solution conditions, particularly pH. Thus, in the case of apoleghemoglobin at least, the equilibrium molten globule formed under destabilizing conditions at acid pH is not a good model for the compact intermediate formed during kinetic refolding experiments. Our high-precision kinetic analysis also reveals an additional slow phase during the folding of apoleghemoglobin, which is not observed for apomyoglobin. Hydrogen exchange pulse-labeling experiments show that the slow-folding phase is associated with residues in the CE loop, which probably forms non-native structure in the intermediate that must be resolved before folding can proceed to completion.
Co-reporter:Gira Bhabha, Lisa Tuttle, Maria A. Martinez-Yamout, Peter E. Wright
FEBS Letters (16 November 2011) Volume 585(Issue 22) pp:3528-3532
Publication Date(Web):16 November 2011
DOI:10.1016/j.febslet.2011.10.014
Dihydrofolate reductase (DHFR) is a well-studied drug target and a paradigm for understanding enzyme catalysis. Preparation of pure DHFR samples, in defined ligand-bound states, is a prerequisite for in vitro studies and drug discovery efforts. We use NMR spectroscopy to monitor ligand content of human and Escherichia coli DHFR (ecDHFR), which bind different co-purifying ligands during expression in bacteria. An alternate purification strategy yields highly pure DHFR complexes, containing only the desired ligands, in the quantities required for structural studies. Interestingly, ecDHFR is bound to endogenous THF while human DHFR is bound to NADP. Consistent with these findings, a designed “humanized” mutant of ecDHFR switches binding specificity in the cell.Highlights► NMR data show that endogenous ligands copurify with DHFR expressed in bacterial cells. ► Human DHFR is preferentially bound to NADP, and Escherichia coli DHFR is bound to THF. ► An E. coli DHFR mutant switches binding specificities to resemble the human enzyme. ► We describe purification schemes to obtain DHFR samples with only desired ligands.
Co-reporter:Rebecca B. Berlow, H. Jane Dyson, Peter E. Wright
FEBS Letters (14 September 2015) Volume 589(Issue 19) pp:2433-2440
Publication Date(Web):14 September 2015
DOI:10.1016/j.febslet.2015.06.003
Intrinsically disordered proteins participate in many important cellular regulatory processes. The absence of a well-defined structure in the free state of a disordered domain, and even on occasion when it is bound to physiological partners, is fundamental to its function. Disordered domains are frequently the location of multiple sites for post-translational modification, the key element of metabolic control in the cell. When a disordered domain folds upon binding to a partner, the resulting complex buries a far greater surface area than in an interaction of comparably-sized folded proteins, thus maximizing specificity at modest protein size. Disorder also maintains accessibility of sites for post-translational modification. Because of their inherent plasticity, disordered domains frequently adopt entirely different structures when bound to different partners, increasing the repertoire of available interactions without the necessity for expression of many different proteins. This feature also adds to the faithfulness of cellular regulation, as the availability of a given disordered domain depends on competition between various partners relevant to different cellular processes.
Co-reporter:Chiaki Nishimura, H. Jane Dyson, Peter E. Wright
Journal of Molecular Biology (12 March 2010) Volume 396(Issue 5) pp:1319-1328
Publication Date(Web):12 March 2010
DOI:10.1016/j.jmb.2009.12.040
Apomyoglobin folds by a sequential mechanism in which the A, G, and H helix regions undergo rapid collapse to form a compact intermediate onto which the central portion of the B helix subsequently docks. To investigate the factors that frustrate folding, we have made mutations in the N-terminus of the B helix to stabilize helical structure (in the mutant G23A/G25A) and to promote native-like hydrophobic packing interactions with helix G (in the mutant H24L/H119F). The kinetic and equilibrium intermediates of G23A/G25A and H24L/H119F were studied by hydrogen exchange pulse labeling and interrupted hydrogen/deuterium exchange combined with NMR. For both mutants, stabilization of helical structure in the N-terminal region of the B helix is confirmed by increased exchange protection in the equilibrium molten globule states near pH 4. Increased protection is also observed in the GH turn region in the G23A/G25A mutant, suggesting that stabilization of the B helix facilitates native-like interactions with the C-terminal region of helix G. These interactions are further enhanced in H24L/H119F. The kinetic burst phase intermediates of both mutants show increased protection, relative to wild-type protein, of amides in the N-terminus of the B helix and in part of the E helix. Stabilization of the E helix in the intermediate is attributed to direct interactions between E helix residues and the newly stabilized N-terminus of helix B. Stabilization of native packing between the B and G helices in H24L/H119F also favors formation of native-like interactions in the GH turn and between the G and H helices in the ensemble of burst phase intermediates. We conclude that instability at the N-terminus of the B helix of apomyoglobin contributes to the energetic frustration of folding by preventing docking and stabilization of the E helix.
Co-reporter:Ariane L. Jansma, Maria A. Martinez-Yamout, Rong Liao, Peiqing Sun, ... Peter E. Wright
Journal of Molecular Biology (12 December 2014) Volume 426(Issue 24) pp:4030-4048
Publication Date(Web):12 December 2014
DOI:10.1016/j.jmb.2014.10.021
•The HPV E7 oncoprotein binds with high affinity to the TAZ2 domain of CBP/p300.•E7 competes with the p53 activation domain for binding to TAZ2.•The full-length E7 dimer promotes association of CBP and pRb in a ternary complex.•The E7 dimer promotes pRb acetylation in cells by a CBP/p300-mediated mechanism.•E7, CBP/p300, and pRb interactions are potentially important for cellular transformation.The oncoprotein E7 from human papillomavirus (HPV) strains that confer high cancer risk mediates cell transformation by deregulating host cellular processes and activating viral gene expression through recruitment of cellular proteins such as the retinoblastoma protein (pRb) and the cyclic-AMP response element binding binding protein (CBP) and its paralog p300. Here we show that the intrinsically disordered N-terminal region of E7 from high-risk HPV16 binds the TAZ2 domain of CBP with greater affinity than E7 from low-risk HPV6b. HPV E7 and the tumor suppressor p53 compete for binding to TAZ2. The TAZ2 binding site in E7 overlaps the LxCxE motif that is crucial for interaction with pRb. While TAZ2 and pRb compete for binding to a monomeric E7 polypeptide, the full-length E7 dimer mediates an interaction between TAZ2 and pRb by promoting formation of a ternary complex. Cell-based assays show that expression of full-length HPV16 E7 promotes increased pRb acetylation and that this response depends both on the presence of CBP/p300 and on the ability of E7 to form a dimer. These observations suggest a model for the oncogenic effect of high-risk HPV16 E7. The disordered region of one E7 molecule in the homodimer interacts with the pocket domain of pRb, while the same region of the other E7 molecule binds the TAZ2 domain of CBP/p300. Through its ability to dimerize, E7 recruits CBP/p300 and pRb into a ternary complex, bringing the histone acetyltransferase domain of CBP/p300 into proximity to pRb and promoting acetylation, leading to disruption of cell cycle control.Download high-res image (72KB)Download full-size image
Co-reporter:Ariane L. Jansma, Maria A. Martinez-Yamout, Rong Liao, Peiqing Sun, ... Peter E. Wright
Journal of Molecular Biology (12 December 2014) Volume 426(Issue 24) pp:4030-4048
Publication Date(Web):12 December 2014
DOI:10.1016/j.jmb.2014.10.021
•The HPV E7 oncoprotein binds with high affinity to the TAZ2 domain of CBP/p300.•E7 competes with the p53 activation domain for binding to TAZ2.•The full-length E7 dimer promotes association of CBP and pRb in a ternary complex.•The E7 dimer promotes pRb acetylation in cells by a CBP/p300-mediated mechanism.•E7, CBP/p300, and pRb interactions are potentially important for cellular transformation.The oncoprotein E7 from human papillomavirus (HPV) strains that confer high cancer risk mediates cell transformation by deregulating host cellular processes and activating viral gene expression through recruitment of cellular proteins such as the retinoblastoma protein (pRb) and the cyclic-AMP response element binding binding protein (CBP) and its paralog p300. Here we show that the intrinsically disordered N-terminal region of E7 from high-risk HPV16 binds the TAZ2 domain of CBP with greater affinity than E7 from low-risk HPV6b. HPV E7 and the tumor suppressor p53 compete for binding to TAZ2. The TAZ2 binding site in E7 overlaps the LxCxE motif that is crucial for interaction with pRb. While TAZ2 and pRb compete for binding to a monomeric E7 polypeptide, the full-length E7 dimer mediates an interaction between TAZ2 and pRb by promoting formation of a ternary complex. Cell-based assays show that expression of full-length HPV16 E7 promotes increased pRb acetylation and that this response depends both on the presence of CBP/p300 and on the ability of E7 to form a dimer. These observations suggest a model for the oncogenic effect of high-risk HPV16 E7. The disordered region of one E7 molecule in the homodimer interacts with the pocket domain of pRb, while the same region of the other E7 molecule binds the TAZ2 domain of CBP/p300. Through its ability to dimerize, E7 recruits CBP/p300 and pRb into a ternary complex, bringing the histone acetyltransferase domain of CBP/p300 into proximity to pRb and promoting acetylation, leading to disruption of cell cycle control.Download high-res image (72KB)Download full-size image
Co-reporter:Peter E. Wright
Journal of Molecular Biology (11 September 2009) Volume 392(Issue 1) pp:1
Publication Date(Web):11 September 2009
DOI:10.1016/j.jmb.2009.07.024
Co-reporter:R. Bryn Fenwick, David Oyen and Peter E. Wright
Physical Chemistry Chemical Physics 2016 - vol. 18(Issue 8) pp:NaN5798-5798
Publication Date(Web):2015/09/24
DOI:10.1039/C5CP04670J
Carr–Purcell–Meiboom–Gill (CPMG) relaxation dispersion measurements are a valuable tool for the characterization of structural transitions on the micro–millisecond timescale. While the measurement of 15N relaxation dispersion is now routine, the measurements with alternative nuclei remain limited. Here we report 15N as well as 1H R2 relaxation dispersion measurements of the N23PP/S148A “dynamic knockout” mutant of dihydrofolate reductase. The 1H dispersion measurements are complementary to 15N data as many additional residues are observed to have dispersive behavior for the 1H nucleus. Simultaneous fitting of the dispersion profiles for the two nuclei increases the accuracy of exchange parameters determined for individual residues and clustered groups of residues. The different sensitivity of the two nuclei to changes in backbone torsional angles, ring currents, and hydrogen bonding effects provides important insights into the nature of the structural changes that take place during the exchange process. We observe clear evidence of direct and indirect hydrogen bond effects for the 15N and 1H chemical shift changes in the active-site, modulation of ring current shielding in the CD-loop and backbone torsional changes in a cluster of residues associated with the C-terminus. This work demonstrates the power of combined 1H and 15N probes for the study of backbone dynamics on the micro-millisecond timescale though the analysis of chemical shift changes.
.jpg)








![Octanamide,3-oxo-N-[(3S)-tetrahydro-2-oxo-3-furanyl]-](http://img.cochemist.com/ccimg/147800/147795-39-9.png)
![Octanamide,3-oxo-N-[(3S)-tetrahydro-2-oxo-3-furanyl]-](http://img.cochemist.com/ccimg/147800/147795-39-9_b.png)




![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-histidyl]-](http://img.cochemist.com/ccimg/115700/115627-72-0.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-histidyl]-](http://img.cochemist.com/ccimg/115700/115627-72-0_b.png)
![L-Valine, N-[N2-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-lysyl]-](http://img.cochemist.com/ccimg/115700/115627-70-8.png)
![L-Valine, N-[N2-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-lysyl]-](http://img.cochemist.com/ccimg/115700/115627-70-8_b.png)
![L-Valine, N-[N2-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-glutaminyl]-](http://img.cochemist.com/ccimg/115700/115627-68-4.png)
![L-Valine, N-[N2-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-glutaminyl]-](http://img.cochemist.com/ccimg/115700/115627-68-4_b.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-seryl]-](http://img.cochemist.com/ccimg/115700/115627-66-2.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-seryl]-](http://img.cochemist.com/ccimg/115700/115627-66-2_b.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-phenylalanyl]-](http://img.cochemist.com/ccimg/115700/115627-64-0.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-phenylalanyl]-](http://img.cochemist.com/ccimg/115700/115627-64-0_b.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-leucyl]-](http://img.cochemist.com/ccimg/115700/115627-62-8.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-leucyl]-](http://img.cochemist.com/ccimg/115700/115627-62-8_b.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-alanyl]-](http://img.cochemist.com/ccimg/115700/115627-60-6.png)
![L-Valine, N-[N-[N-(1-L-tyrosyl-L-prolyl)-L-tyrosyl]-L-alanyl]-](http://img.cochemist.com/ccimg/115700/115627-60-6_b.png)


![3,5,9-Trioxa-4-phosphaheneicosan-1-aminium,4-hydroxy-N,N,N-trimethyl-10-oxo-7-[(1-oxododecyl)oxy]-, inner salt, 4-oxide,(7R)-](http://img.cochemist.com/ccimg/18200/18194-25-7.png)
![3,5,9-Trioxa-4-phosphaheneicosan-1-aminium,4-hydroxy-N,N,N-trimethyl-10-oxo-7-[(1-oxododecyl)oxy]-, inner salt, 4-oxide,(7R)-](http://img.cochemist.com/ccimg/18200/18194-25-7_b.png)



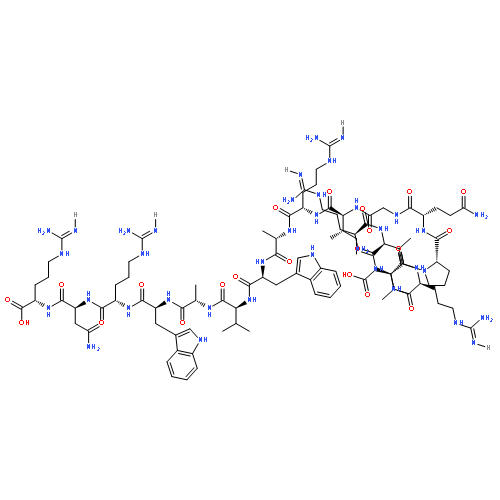



![1,5-Cyclohexadiene-1-carboxylicacid, 3-[(1-carboxyethenyl)oxy]-4-hydroxy-, (3R,4R)-](http://img.cochemist.com/ccimg/700/617-12-9.png)
![1,5-Cyclohexadiene-1-carboxylicacid, 3-[(1-carboxyethenyl)oxy]-4-hydroxy-, (3R,4R)-](http://img.cochemist.com/ccimg/700/617-12-9_b.png)








![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)








![L-Glutamic acid,N-[4-[[(2-amino-3,4,7,8-tetrahydro-4-oxo-6-pteridinyl)methyl]amino]benzoyl]-](http://img.cochemist.com/ccimg/4100/4033-27-6.png)
![L-Glutamic acid,N-[4-[[(2-amino-3,4,7,8-tetrahydro-4-oxo-6-pteridinyl)methyl]amino]benzoyl]-](http://img.cochemist.com/ccimg/4100/4033-27-6_b.png)




![L-Glutamic acid,N-[4-[2-(2-amino-3,4,5,6,7,8-hexahydro-4-oxopyrido[2,3-d]pyrimidin-6-yl)ethyl]benzoyl]-](http://img.cochemist.com/ccimg/95700/95693-76-8.png)
![L-Glutamic acid,N-[4-[2-(2-amino-3,4,5,6,7,8-hexahydro-4-oxopyrido[2,3-d]pyrimidin-6-yl)ethyl]benzoyl]-](http://img.cochemist.com/ccimg/95700/95693-76-8_b.png)


![Glutamic acid,N-[4-[[(2-amino-3,4,5,6,7,8-hexahydro-4-oxo-6-pteridinyl)methyl]amino]benzoyl]-](http://img.cochemist.com/ccimg/29400/29347-89-5.png)
![Glutamic acid,N-[4-[[(2-amino-3,4,5,6,7,8-hexahydro-4-oxo-6-pteridinyl)methyl]amino]benzoyl]-](http://img.cochemist.com/ccimg/29400/29347-89-5_b.png)

