Co-reporter:Robert W. Walters;Tyler Matheny;Laura S. Mizoue;Bhalchandra S. Rao;Denise Muhlrad
PNAS 2017 Volume 114 (Issue 3 ) pp:480-485
Publication Date(Web):2017-01-17
DOI:10.1073/pnas.1619369114
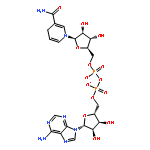
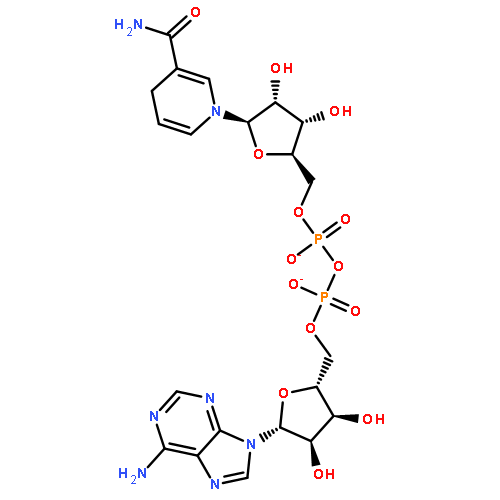
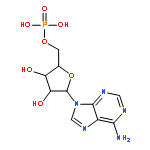
RNAs besides tRNA and rRNA contain chemical modifications, including the recently described 5′ nicotinamide-adenine dinucleotide
(NAD+) RNA in bacteria. Whether 5′ NAD-RNA exists in eukaryotes remains unknown. We demonstrate that 5′ NAD-RNA is found on subsets
of nuclear and mitochondrial encoded mRNAs in Saccharomyces cerevisiae. NAD-mRNA appears to be produced cotranscriptionally because NAD-RNA is also found on pre-mRNAs, and only on mitochondrial
transcripts that are not 5′ end processed. These results define an additional 5′ RNA cap structure in eukaryotes and raise
the possibility that this 5′ NAD+ cap could modulate RNA stability and translation on specific subclasses of mRNAs.
Co-reporter:Joshua R. Wheeler, Saumya Jain, Anthony Khong, Roy Parker
Methods 2017 Volume 126(Volume 126) pp:
Publication Date(Web):15 August 2017
DOI:10.1016/j.ymeth.2017.04.020
•Stress granule cores are stable ex vivo.•Stress granule cores contain known stress granule components and RNA.•Stress granule cores are purified using a combination of centrifugation followed by immunoprecipitation.•A purified population of stress granule cores can be used for biochemical, proteomic, and transcriptomic experiments.Stress granules are dynamic, conserved RNA-protein (RNP) assemblies that form when translation is limiting; and are related to pathological aggregates in degenerative disease. Mammalian stress granules are comprised of two structures – an unstable shell and more stable cores. Herein we describe methodology for isolation of stress granule cores from both yeast and mammalian cells. The protocol consists of first enriching for stress granule cores using centrifugation and then further purifying stress granule cores using immunoprecipitation. The stress granule core isolation protocol provides a starting point for assisting future endeavors aimed at discovering conserved RNA regulatory mechanisms and potential links between RNP aggregation and degenerative disease.
Co-reporter:Anthony Khong, Tyler Matheny, Saumya Jain, Sarah F. Mitchell, ... Roy Parker
Molecular Cell 2017 Volume 68, Issue 4(Volume 68, Issue 4) pp:
Publication Date(Web):16 November 2017
DOI:10.1016/j.molcel.2017.10.015
•Purification of stress granule cores identifies RNAs in stress granules•Only ∼10% of bulk mRNA molecules are present in stress granules•Most mRNAs go to stress granules, but the efficiency varies from <1% to >95%•Poor translation efficiency and longer length correlate with targeting to granulesStress granules are mRNA-protein assemblies formed from nontranslating mRNAs. Stress granules are important in the stress response and may contribute to some degenerative diseases. Here, we describe the stress granule transcriptome of yeast and mammalian cells through RNA-sequencing (RNA-seq) analysis of purified stress granule cores and single-molecule fluorescence in situ hybridization (smFISH) validation. While essentially every mRNA, and some noncoding RNAs (ncRNAs), can be targeted to stress granules, the targeting efficiency varies from <1% to >95%. mRNA accumulation in stress granules correlates with longer coding and UTR regions and poor translatability. Quantifying the RNA-seq analysis by smFISH reveals that only 10% of bulk mRNA molecules accumulate in mammalian stress granules and that only 185 genes have more than 50% of their mRNA molecules in stress granules. These results suggest that stress granules may not represent a specific biological program of messenger ribonucleoprotein (mRNP) assembly, but instead form by condensation of nontranslating mRNPs in proportion to their length and lack of association with ribosomes.Download high-res image (272KB)Download full-size image
Co-reporter:Siddharth Shukla
PNAS 2014 111 (32 ) pp:E3277-E3286
Publication Date(Web):2014-08-12
DOI:10.1073/pnas.1412614111
The accurate biogenesis of RNA–protein complexes is a key aspect of eukaryotic cells. Defects in Sm protein complex binding
to snRNAs are known to reduce levels of snRNAs, suggesting an unknown quality control system for small nuclear ribonucleoprotein
(snRNP) assembly. snRNA quality control may also be relevant in spinal muscular atrophy, which is caused by defects in the
survival motor neuron (SMN)1 gene, an assembly factor for loading the Sm complex on snRNAs and, when severely reduced, can lead to reduced levels of snRNAs
and splicing defects. To determine how assembly-defective snRNAs are degraded, we first demonstrate that yeast U1 Sm-mutant
snRNAs are degraded either by Rrp6- or by Dcp2-dependent decapping/5′-to-3′ decay. Knockdown of the decapping enzyme DCP2
in mammalian cells also increases the levels of assembly-defective snRNAs and suppresses some splicing defects seen in SMN-deficient
cells. These results identify a conserved mechanism of snRNA quality control, and also suggest a general paradigm wherein
the phenotype of an “RNP assembly disease” might be suppressed by inhibition of a competing RNA quality control mechanism.
Co-reporter:Yuriko Harigaya
PNAS 2012 Volume 109 (Issue 29 ) pp:11764-11769
Publication Date(Web):2012-07-17
DOI:10.1073/pnas.1119741109
The general pathways of eukaryotic mRNA decay occur via deadenylation followed by 3′ to 5′ degradation or decapping, although
some endonuclease sites have been identified in metazoan mRNAs. To determine the role of endonucleases in mRNA degradation
in Saccharomyces cerevisiae, we mapped 5′ monophosphate ends on mRNAs in wild-type and dcp2∆ xrn1∆ yeast cells, wherein mRNA endonuclease cleavage products are stabilized. This led to three important observations. First,
only few mRNAs that undergo low-level endonucleolytic cleavage were observed, suggesting that endonucleases are not a major
contributor to yeast mRNA decay. Second, independent of known decapping enzymes, we observed low levels of 5′ monophosphates
on some mRNAs, suggesting that an unknown mechanism can generate 5′ exposed ends, although for all substrates tested, Dcp2
was the primary decapping enzyme. Finally, we identified debranched lariat intermediates from intron-containing genes, demonstrating
a significant discard pathway for mRNAs during the second step of pre-mRNA splicing, which is a potential step to regulate
gene expression.
Co-reporter:David S.W. Protter, Roy Parker
Trends in Cell Biology (September 2016) Volume 26(Issue 9) pp:668-679
Publication Date(Web):1 September 2016
DOI:10.1016/j.tcb.2016.05.004
Stress granules are assemblies of untranslating messenger ribonucleoproteins (mRNPs) that form from mRNAs stalled in translation initiation. Stress granules form through interactions between mRNA-binding proteins that link together populations of mRNPs. Interactions promoting stress granule formation include conventional protein–protein interactions as well as interactions involving intrinsically disordered regions (IDRs) of proteins. Assembly and disassembly of stress granules are modulated by various post-translational modifications as well as numerous ATP-dependent RNP or protein remodeling complexes, illustrating that stress granules represent an active liquid wherein energy input maintains their dynamic state. Stress granule formation modulates the stress response, viral infection, and signaling pathways. Persistent or aberrant stress granule formation contributes to neurodegenerative disease and some cancers.
Co-reporter:Siddharth Shukla, Roy Parker
Trends in Molecular Medicine (July 2016) Volume 22(Issue 7) pp:615-628
Publication Date(Web):1 July 2016
DOI:10.1016/j.molmed.2016.05.005
A key aspect of cellular function is the proper assembly and utilization of ribonucleoproteins (RNPs). Recent studies have shown that hyper- or hypo-assembly of various RNPs can lead to human diseases. Defects in the formation of RNPs lead to ‘RNP hypo-assembly diseases’, which can be caused by RNA degradation outcompeting RNP assembly. By contrast, excess RNP assembly, either in higher order RNP granules, or due to the expression of repeat-containing RNAs, can lead to ‘RNP hyper-assembly diseases’. Here, we discuss the most recent advances in understanding the cause of disease onset, as well as potential therapies from the aspect of modulating RNP assembly in the cell, which presents a novel route to the treatment of these diseases.
Co-reporter:Yuriko Harigaya, Roy Parker
Molecular Cell (8 May 2014) Volume 54(Issue 3) pp:330-332
Publication Date(Web):8 May 2014
DOI:10.1016/j.molcel.2014.04.027
In this issue of Molecular Cell, Chen et al. (2014) provide evidence that FMRP represses translation by binding the ribosome, suggesting a novel form of translational control.
Co-reporter:Sarah F. Mitchell, Roy Parker
Molecular Cell (22 May 2014) Volume 54(Issue 4) pp:547-558
Publication Date(Web):22 May 2014
DOI:10.1016/j.molcel.2014.04.033
The proper processing, export, localization, translation, and degradation of mRNAs are necessary for regulation of gene expression. These processes are controlled by mRNA-specific regulatory proteins, noncoding RNAs, and core machineries common to most mRNAs. These factors bind the mRNA in large complexes known as messenger ribonucleoprotein particles (mRNPs). Herein, we review the components of mRNPs, how they assemble and rearrange, and how mRNP composition differentially affects mRNA biogenesis, function, and degradation. We also describe how properties of the mRNP “interactome” lead to emergent principles affecting the control of gene expression.
Co-reporter:Yuan Lin, David S.W. Protter, Michael K. Rosen, Roy Parker
Molecular Cell (15 October 2015) Volume 60(Issue 2) pp:208-219
Publication Date(Web):15 October 2015
DOI:10.1016/j.molcel.2015.08.018
•Intrinsically disordered regions (IDRs) of RNA-binding proteins can phase separate•RNA, crowding agents, and low salt promote IDR phase separation•Phase-separated droplets stabilize over time by formation of amyloid-like fibers•Multiple IDRs can be recruited to phase-separated dropletsEukaryotic cells possess numerous dynamic membrane-less organelles, RNP granules, enriched in RNA and RNA-binding proteins containing disordered regions. We demonstrate that the disordered regions of key RNP granule components and the full-length granule protein hnRNPA1 can phase separate in vitro, producing dynamic liquid droplets. Phase separation is promoted by low salt concentrations or RNA. Over time, the droplets mature to more stable states, as assessed by slowed fluorescence recovery after photobleaching and resistance to salt. Maturation often coincides with formation of fibrous structures. Different disordered domains can co-assemble into phase-separated droplets. These biophysical properties demonstrate a plausible mechanism by which interactions between disordered regions, coupled with RNA binding, could contribute to RNP granule assembly in vivo through promoting phase separation. Progression from dynamic liquids to stable fibers may be regulated to produce cellular structures with diverse physiochemical properties and functions. Misregulation could contribute to diseases involving aberrant RNA granules.Download high-res image (209KB)Download full-size image
.png)