Co-reporter:Andrew Welland, Pierre E. Garnaud, Maki Kitamura, Caroline S. Miles and Simon Daff
Biochemistry 2008 Volume 47(Issue 37) pp:
Publication Date(Web):August 22, 2008
DOI:10.1021/bi800787m
Calmodulin (CaM) activates NO synthase (NOS) by binding to a 20 amino acid interdomain hinge in the presence of Ca2+, inducing electrons to be transferred from the FAD to the heme of the enzyme via a mobile FMN domain. The activation process is influenced by a number of structural features, including an autoinhibitory loop, the C-terminal tail of the enzyme, and a number of phosphorylation sites. Crystallographic and other recent experimental data imply that the regulatory elements lie within the interface between the FAD- and FMN-binding domains, restricting the movement of the two cofactors with respect to each other. Arg1229 of rat neuronal NOS is a conserved residue in the FAD domain that forms one of only two electrostatic contacts between the domains. Mutation of this residue to Glu reverses its charge and is expected to induce an interdomain repulsion, allowing the importance of the interface and domain−domain motion to be probed. The charge-reversal mutation R1229E has three dramatic effects on catalysis: (i) hydride transfer from NADPH to FAD is activated in the CaM-free enzyme, (ii) FAD to FMN electron transfer is inhibited in both forms, and (iii) electron transfer from FMN to the surrogate acceptor cytochrome c is activated in the CaM-free enzyme. As a result, during steady-state turnover with cytochrome c, calmodulin now deactivates the enzyme and causes cytochrome c-dependent inhibition. Evidently, domain−domain separation is large enough in the mutant to accommodate another protein between the cofactors. The effects of this single charge reversal on three distinct catalytic events illustrate how each is differentially dependent on the enzyme conformation and support a model for catalytic motion in which steps i, ii, and iii occur in the hinged open, closed, and open states, respectively. This model is also likely to apply to related enzymes such as cytochrome P450 reductase.
Co-reporter:Tobias W.B. Ost, Sidong Liu, Simon Daff
Nitric Oxide (1 August 2011) Volume 25(Issue 2) pp:89-94
Publication Date(Web):1 August 2011
DOI:10.1016/j.niox.2011.01.008
Nitric oxide is known to coordinate to ferrous heme proteins very tightly, following which it is susceptible to reaction with molecular oxygen or free NO. Its coordination to ferric heme is generally weaker but the resultant complexes are more stable in the presence of oxygen. Here we report determination of the binding constants of Cytochrome P450 BM3 for nitric oxide in the ferric state in the presence and absence of substrate. Compared to other 5-coordinate heme proteins, the Kd values are particularly low at 16 and 40 nM in the presence and absence of substrate respectively. This most likely reflects the high hydrophobicity of the active site of this enzyme. The binding of NO is tight enough to enable P450 BM3 oxygenase domain to be used to determine NO concentrations and in real-time NO detection assays, which would be particularly useful under conditions of low oxygen concentration, where current methods break down.
Co-reporter:Simon Daff
Nitric Oxide (1 August 2010) Volume 23(Issue 1) pp:1-11
Publication Date(Web):1 August 2010
DOI:10.1016/j.niox.2010.03.001
Production of NO from arginine and molecular oxygen is a complex chemical reaction unique to biology. Our understanding of the chemical and regulation mechanisms of the NO synthases has developed over the past two decades, uncovering some extraordinary features. This article reviews recent progress and highlights current issues and controversies. The structure of the enzyme has now been determined almost in entirety, although it is as a selection of fragments, which are difficult to assemble unambiguously. NO synthesis is driven by electron transfer through FAD and FMN cofactors, which is controlled by calmodulin binding in the constitutive mammalian enzymes. Many of the unique structural features involved have been characterised, but the mechanics of calmodulin-dependent activation are largely unresolved. Ultimately, NO is produced in the active site by the reaction of arginine with activated heme-bound oxygen in two distinct cycles. The unique role of the tetrahydrobiopterin cofactor as an electron donor in this process has now been established, but the subsequent chemical events are currently a matter of intense speculation and debate.

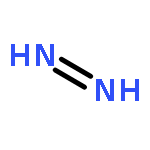
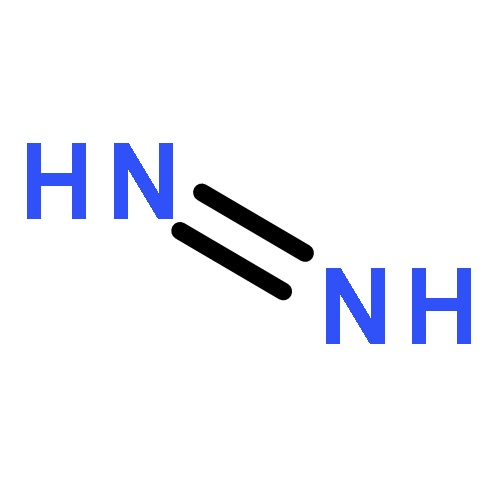






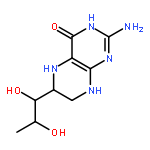

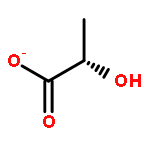
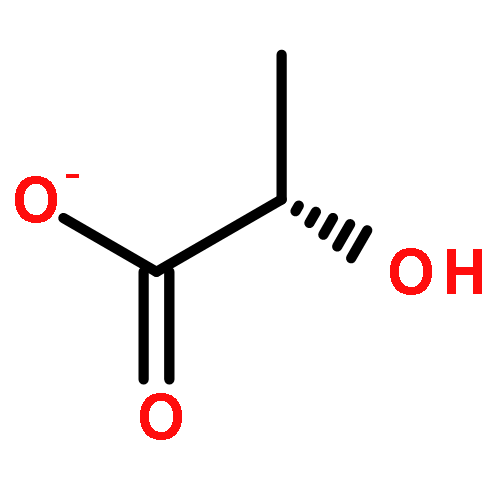
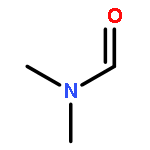
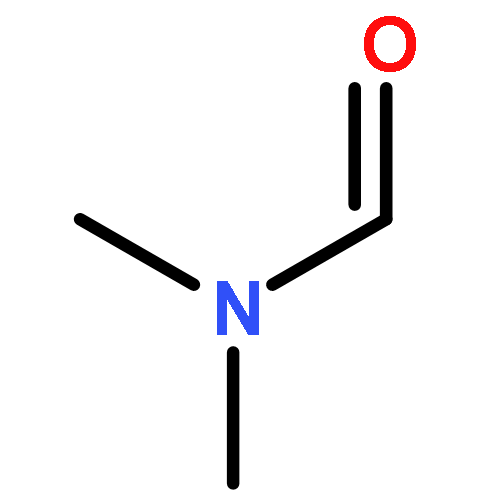
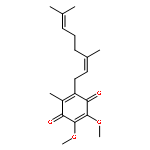

![L-Ornithine,N5-[(hydroxyamino)iminomethyl]-](http://img.cochemist.com/ccimg/53100/53054-07-2.png)
![L-Ornithine,N5-[(hydroxyamino)iminomethyl]-](http://img.cochemist.com/ccimg/53100/53054-07-2_b.png)


![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]nitrosyl-, hydrogen (1:2), (SP-5-13)-](/data/chemimg/1130700/54854-55-6.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]nitrosyl-, hydrogen (1:2), (SP-5-13)-](/data/chemimg/1130700/54854-55-6_b.png)






![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)









