Co-reporter:Nathaniel S. Sickerman, Kazuki TanifujiChi Chung Lee, Yasuhiro Ohki, Kazuyuki Tatsumi, Markus W. Ribbe, Yilin Hu
Journal of the American Chemical Society 2017 Volume 139(Issue 2) pp:603-606
Publication Date(Web):January 3, 2017
DOI:10.1021/jacs.6b11633
Solvent-extracted nitrogenase cofactors can reduce C1 substrates (CN–, CO and CO2) to hydrocarbons in reactions driven by a strong reductant, SmI2 (E0′ = −1.55 V vs SCE). Here we show that a synthetic [Et4N]4[Fe6S9(SEt)2] cluster (designated the Fe6RHH-cluster), which mimics the homometallic [Fe8S9C] core of the nitrogenase cofactor (designated the L-cluster), is capable of conversion of C1 substrates into hydrocarbons in the same reactions. Comparison of the yields and product profiles between these homometallic clusters and their heterometallic counterparts points to possible roles of the heterometal, interstitial carbide and belt sulfur-bridged iron atoms in catalysis. More importantly, the observation that a “simplified”, homometallic cofactor mimic can perform Fischer–Tropsch-like hydrocarbon synthesis suggests future biotechnological adaptability of nitrogenase-based biomimetic compounds for recycling C1 substrates into useful chemical and fuel products.
Co-reporter:Yilin Hu, Markus W Ribbe
Current Opinion in Chemical Biology 2016 Volume 31() pp:188-194
Publication Date(Web):April 2016
DOI:10.1016/j.cbpa.2016.02.016
•The carbide of nitrogense M-cluster originates from the methyl group of SAM.•The insertion of carbide is catalyzed by NifB via a radical SAM-dependent mechanism.•The methyl group is transferred from SAM to an sulfur atom of the M-cluster precursor.•The transfer of methyl to the M-cluster precursor occurs before hydrogen abstraction.•The NifB protein family represents as a new class of radical SAM methyltransferases.Nitrogenase catalyzes the important reactions of N2-reduction, CO-reduction and CO2-reduction at its active cofactor site. Designated the M-cluster, this complex metallocofactor is assembled through the generation of a characteristic 8Fe-core before the insertion of Mo and homocitrate that completes the stoichiometry of the M-cluster. NifB catalyzes the crucial step of radical SAM-dependent carbide insertion that occurs concomitant with the insertion a ‘9th’ sulfur and the rearrangement/coupling of two 4Fe-clusters into a complete 8Fe-core of the M-cluster. Further categorization of a family of NifB proteins as a new class of radical SAM methyltransferases suggests a general function of these proteins in complex metallocofactor assembly and provides a new platform for unveiling unprecedented chemical reactions catalyzed by biological systems.
Co-reporter:Aaron W. Fay;Chi Chung Lee;Michael A. Blank;Johannes G. Rebelein;Britt Hedman;Keith O. Hodgson;Yilin Hu
PNAS 2016 Volume 113 (Issue 34 ) pp:9504-9508
Publication Date(Web):2016-08-23
DOI:10.1073/pnas.1609574113
NifEN is a biosynthetic scaffold for the cofactor of Mo-nitrogenase (designated the M-cluster). Previous studies have revealed
the sequence and structural homology between NifEN and NifDK, the catalytic component of nitrogenase. However, direct proof
for the functional homology between the two proteins has remained elusive. Here we show that, upon maturation of a cofactor
precursor (designated the L-cluster) on NifEN, the cluster species extracted from NifEN is spectroscopically equivalent and
functionally interchangeable with the native M-cluster extracted from NifDK. Both extracted clusters display nearly indistinguishable
EPR features, X-ray absorption spectroscopy/extended X-ray absorption fine structure (XAS/EXAFS) spectra and reconstitution
activities, firmly establishing the M-cluster–bound NifEN (designated NifENM) as the only protein other than NifDK to house the unique nitrogenase cofactor. Iron chelation experiments demonstrate a
relocation of the cluster from the surface to its binding site within NifENM upon maturation, which parallels the insertion of M-cluster into an analogous binding site in NifDK, whereas metal analyses
suggest an asymmetric conformation of NifENM with an M-cluster in one αβ-half and an empty cluster-binding site in the other αβ-half, which led to the proposal of a stepwise
assembly mechanism of the M-cluster in the two αβ-dimers of NifEN. Perhaps most importantly, NifENM displays comparable ATP-independent substrate-reducing profiles to those of NifDK, which establishes the M-cluster–bound
αβ-dimer of NifENM as a structural and functional mimic of one catalytic αβ-half of NifDK while suggesting the potential of this protein as
a useful tool for further investigations of the mechanistic details of nitrogenase.
Co-reporter:Chi Chung Lee;Aaron W. Fay;Tsu-Chien Weng;Courtney M. Krest;Britt Hedman;Keith O. Hodgson;Yilin Hu
PNAS 2015 Volume 112 (Issue 45 ) pp:13845-13849
Publication Date(Web):2015-11-10
DOI:10.1073/pnas.1519696112
Biocatalysis by nitrogenase, particularly the reduction of N2 and CO by this enzyme, has tremendous significance in environment- and energy-related areas. Elucidation of the detailed
mechanism of nitrogenase has been hampered by the inability to trap substrates or intermediates in a well-defined state. Here,
we report the capture of substrate CO on the resting-state vanadium-nitrogenase in a catalytically competent conformation.
The close resemblance of this active CO-bound conformation to the recently described structure of CO-inhibited molybdenum-nitrogenase
points to the mechanistic relevance of sulfur displacement to the activation of iron sites in the cofactor for CO binding.
Moreover, the ability of vanadium-nitrogenase to bind substrate in the resting-state uncouples substrate binding from subsequent
turnover, providing a platform for generation of defined intermediate(s) of both CO and N2 reduction.
Co-reporter:Dr. Chi Chung Lee;Dr. Yilin Hu;Dr. Markus W. Ribbe
Angewandte Chemie International Edition 2015 Volume 54( Issue 4) pp:1219-1222
Publication Date(Web):
DOI:10.1002/anie.201410412
Abstract
Nitrogenase cofactors can be extracted into an organic solvent to catalyze the reduction of cyanide (CN−), carbon monoxide (CO), and carbon dioxide (CO2) without using adenosine triphosphate (ATP), when samarium(II) iodide (SmI2) and 2,6-lutidinium triflate (Lut-H) are employed as a reductant and a proton source, respectively. Driven by SmI2, the cofactors catalytically reduce CN− or CO to C1–C4 hydrocarbons, and CO2 to CO and C1–C3 hydrocarbons. The CC coupling from CO2 indicates a unique Fischer–Tropsch-like reaction with an atypical carbonaceous substrate, whereas the catalytic turnover of CN−, CO, and CO2 by isolated cofactors suggests the possibility to develop nitrogenase-based electrocatalysts for the production of hydrocarbons from these carbon-containing compounds.
Co-reporter:Dr. Markus W. Ribbe
Angewandte Chemie International Edition 2015 Volume 54( Issue 29) pp:8337-8339
Publication Date(Web):
DOI:10.1002/anie.201503979
Co-reporter:Dr. Markus W. Ribbe
Angewandte Chemie 2015 Volume 127( Issue 29) pp:8455-8457
Publication Date(Web):
DOI:10.1002/ange.201503979
Co-reporter:Markus W. Ribbe, Yilin Hu, Keith O. Hodgson, and Britt Hedman
Chemical Reviews 2014 Volume 114(Issue 8) pp:4063
Publication Date(Web):December 13, 2013
DOI:10.1021/cr400463x
Co-reporter:Kresimir Rupnik, Chi Chung Lee, Jared A. Wiig, Yilin Hu, Markus W. Ribbe, and Brian J. Hales
Biochemistry 2014 Volume 53(Issue 7) pp:
Publication Date(Web):February 12, 2014
DOI:10.1021/bi401699u
The P-cluster in the nitrogenase MoFe protein is a [Fe8S7] cluster and represents the most complex FeS cluster found in Nature. To date, the exact mechanism of the in vivo synthesis of the P-cluster remains unclear. What is known is that the precursor to the P-cluster is a pair of neighboring [Fe4S4]-like clusters found on the ΔnifH MoFe protein, a protein expressed in the absence of the nitrogenase Fe protein (NifH). Moreover, incubation of the ΔnifH MoFe protein with NifH and MgATP results in the synthesis of the MoFe protein P-clusters. To improve our understanding of the mechanism of this reaction, we conducted a magnetic circular dichroism (MCD) spectroscopic study of the [Fe4S4]-like clusters on the ΔnifH MoFe protein. Reducing the ΔnifH MoFe protein with Ti(III) citrate results in the quenching of the S = 1/2 electron paramagnetic resonance signal associated with the [Fe4S4]+ state of the clusters. MCD spectroscopy reveals this reduction results in all four 4Fe clusters being converted into the unusual, all-ferrous [Fe4S4]0 state. Subsequent increases of the redox potential generate new clusters. Most significantly, one of these newly formed clusters is the P-cluster, which represents approximately 20–25% of the converted Fe concentration. The other two clusters are an X cluster, of unknown structure, and a classic [Fe4S4] cluster, which represents approximately 30–35% of the Fe concentration. Diamagnetic FeS clusters may also have been generated but, because of their low spectral intensity, would not have been identified. These results demonstrate that the nitrogenase P-cluster can be generated in the absence of NifH and MgATP.
Co-reporter:Johannes G. Rebelein;Dr. Yilin Hu;Dr. Markus W. Ribbe
Angewandte Chemie International Edition 2014 Volume 53( Issue 43) pp:11543-11546
Publication Date(Web):
DOI:10.1002/anie.201406863
Abstract
The molybdenum and vanadium nitrogenases are two homologous enzymes with distinct structural and catalytic features. Previously, it was demonstrated that the V nitrogenase was nearly 700 times more active than its Mo counterpart in reducing CO to hydrocarbons. Herein, a similar discrepancy between the two nitrogenases in the reduction of CO2 is reported, with the V nitrogenase being capable of reducing CO2 to CO, CD4, C2D4, and C2D6, and its Mo counterpart only capable of reducing CO2 to CO. Furthermore, it is shown that the V nitrogenase may direct the formation of CD4 in part via CO2-derived CO, but that it does not catalyze the formation of C2D4 and C2D6 along this route. The exciting observation of a V nitrogenase-catalyzed CC coupling with CO2 as the origin of the building blocks adds another interesting reaction to the catalytic repertoire of this unique enzyme system. The differential activities of the V and Mo nitrogenases in CO2 reduction provide an important framework for systematic investigations of this reaction in the future.
Co-reporter:Yilin Hu
JBIC Journal of Biological Inorganic Chemistry 2014 Volume 19( Issue 6) pp:731-736
Publication Date(Web):2014 August
DOI:10.1007/s00775-014-1137-2
Nitrogenase catalyzes the reduction of N2 to NH3, a key step in the global nitrogen cycle. This article describes our journey toward the definition of a complete molecular structure of the active site of nitrogenase, with an emphasis on the discovery of the interstitial carbide and the radical SAM-dependent insertion of this atom into the active FeMo cofactor site of nitrogenase.
Co-reporter:Jared A. Wiig ; Chi Chung Lee ; Yilin Hu
Journal of the American Chemical Society 2013 Volume 135(Issue 13) pp:4982-4983
Publication Date(Web):March 20, 2013
DOI:10.1021/ja401698d
The fate of the interstitial atom of the nitrogenase cofactor during substrate turnover has remained a topic of interest since the discovery of this atom more than a decade ago. In this study, we labeled the interstitial carbide atom with 14C and 13C isotopes and traced the fate of the isotope under turnover conditions. Our results show that the interstitial carbide cannot be exchanged upon turnover, nor can it be used as a substrate and incorporated into the products. These observations point to a role of the interstitial carbide in stabilizing the cofactor structure, although a function of this atom in indirectly modulating the reactivity of the cofactor or directly interacting with the substrate cannot be excluded.
Co-reporter:Kresimir Rupnik ; Yilin Hu ; Chi Chung Lee ; Jared A. Wiig ; Markus W. Ribbe ;Brian J. Hales
Journal of the American Chemical Society 2012 Volume 134(Issue 33) pp:13749-13754
Publication Date(Web):July 28, 2012
DOI:10.1021/ja304077h
Mo nitrogenase consists of two component proteins: the Fe protein, which contains a [Fe4S4] cluster, and the MoFe protein, which contains two different classes of metal cluster: P-cluster ([Fe8S7]) and FeMoco ([MoFe7S9C·homocitrate]). The P-cluster is believed to mediate the electron transfer between the Fe protein and the MoFe protein via interconversions between its various oxidation states, such as the all-ferrous state (PN) and the one- (P+) and two-electron (P2+) oxidized states. While the structural and electronic properties of PN and P2+ states have been well characterized, little is known about the electronic structure of the P+ state. Here, a mutant strain of Azotobacter vinelandii (DJ1193) was used to facilitate the characterization of the P+ state of P-cluster. This strain expresses a MoFe protein variant (designated ΔnifB β-188Cys MoFe protein) that accumulates the P+ form of P-cluster in the resting state. Magnetic circular dichroism (MCD) spectrum of the P-cluster in the oxidized ΔnifB β-188Cys MoFe protein closely resembles that of the P2+ state in the oxidized wild-type MoFe protein, except for the absence of a major charge-transfer band centered at 823 nm. Moreover, magnetization curves of ΔnifB β-188Cys and wild-type MoFe proteins suggest that the P2+ species in both proteins have the same spin state. MCD spectrum of the P+ state in the ΔnifB β-188Cys MoFe protein, on the other hand, is associated with a classic [Fe4S4]+ cluster, suggesting that the P-cluster could be viewed as two coupled 4Fe clusters and that it could donate either one or two electrons to FeMoco by using one or both of its 4Fe halves. Such a mode of action of P-cluster could provide energetic and kinetic advantages to nitrogenase in the complex mechanism of N2 reduction.
Co-reporter:Kyle M. Lancaster ; Yilin Hu ; Uwe Bergmann ; Markus W. Ribbe ;Serena DeBeer
Journal of the American Chemical Society 2012 Volume 135(Issue 2) pp:610-612
Publication Date(Web):December 31, 2012
DOI:10.1021/ja309254g
The iron–molybdenum cofactor (FeMoco) of nitrogenase contains a biologically unprecedented μ6-coordinated C4– ion. Although the role of this interstitial atom in nitrogenase catalysis is unknown, progress in understanding its biosynthetic origins has been made. Here we report valence-to-core Fe Kβ X-ray emission spectroscopy data to show that this C4– ion is present in the Fe8S9 “L-cluster,” which is the immediate precursor to FeMoco prior to the insertion of molybdenum and coordination by homocitrate. These results accord with recent evidence supporting a role for the S-adenosylmethionine-dependent enzyme NifB in the incorporation of carbon into the FeMoco center of nitrogenase.
Co-reporter:Yilin Hu, Chi Chung Lee and Markus W. Ribbe
Dalton Transactions 2012 vol. 41(Issue 4) pp:1118-1127
Publication Date(Web):18 Nov 2011
DOI:10.1039/C1DT11535A
Nitrogenase catalyzes the biological conversion of atmospheric dinitrogen to bioavailable ammonia. The molybdenum (Mo)- and vanadium (V)-dependent nitrogenases are two homologous members of this metalloenzyme family. However, despite their similarities in structure and function, the characterization of V-nitrogenase has taken a much longer and more winding path than that of its Mo-counterpart. From the initial discovery of this nitrogen-fixing system, to the recent finding of its CO-reducing capacity, V-nitrogenase has proven to be a two-hit wonder in the over-a-century-long research of nitrogen fixation. This perspective provides a brief account of the catalytic function and structural basis of V-nitrogenase, as well as a short discussion of the theoretical and practical potentials of this unique metalloenzyme.
Co-reporter:Chi Chung Lee;Dr. Yilin Hu;Dr. Markus W. Ribbe
Angewandte Chemie International Edition 2012 Volume 51( Issue 8) pp:1947-1949
Publication Date(Web):
DOI:10.1002/anie.201108916
Co-reporter:Chi Chung Lee;Yilin Hu
PNAS 2012 Volume 109 (Issue 18 ) pp:6922-6926
Publication Date(Web):2012-05-01
DOI:10.1073/pnas.1202429109
The P-cluster of nitrogenase is largely known for its function to mediate electron transfer to the active cofactor site during
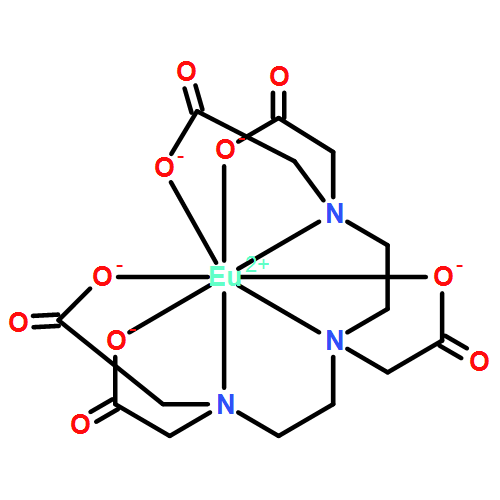
catalysis. Here, we show that a P-cluster variant (designated P*-cluster), which consists of paired [Fe4S4]-like clusters, can catalyze ATP-independent substrate reduction in the presence of a strong reductant, europium(II) diethylenetriaminepentaacetate
[Eu(II)-DTPA]. The observation of a decrease of activity in the rank ΔnifH, ΔnifBΔnifZ, and ΔnifB MoFe protein, which corresponds to a decrease of the amount of P*-clusters in these cofactor-deficient proteins, firmly establishes
P*-cluster as a catalytically active metal center in Eu(II)-DTPA–driven reactions. More excitingly, the fact that P*-cluster
is not only capable of catalyzing the two-electron reduction of proton, acetylene, ethylene, and hydrazine, but also capable
of reducing cyanide, carbon monoxide, and carbon dioxide to alkanes and alkenes, points to a possibility of developing biomimetic
catalysts for hydrocarbon production under ambient conditions.
Co-reporter:Kyle M. Lancaster;Michael Roemelt;Patrick Ettenhuber;Yilin Hu;Frank Neese;Uwe Bergmann;Serena DeBeer
Science 2011 Vol 334(6058) pp:974-977
Publication Date(Web):18 Nov 2011
DOI:10.1126/science.1206445
A central light atom in a cofactor at the nitrogenase active site is identified as a carbon.
Co-reporter:Yilin Hu, Markus W. Ribbe
Coordination Chemistry Reviews 2011 Volume 255(9–10) pp:1218-1224
Publication Date(Web):May 2011
DOI:10.1016/j.ccr.2010.11.018
Biosynthesis of nitrogenase FeMoco is a highly complex process that requires, minimally, the participation of nifS, nifU, nifB, nifE, nifN, nifV, nifH, nifD and nifK gene products. Previous genetic analyses have identified the essential factors for the assembly of FeMoco; however, the exact functions of these factors and the precise sequence of events during the assembly process had remained unclear until recently, when a number of the biosynthetic intermediates of FeMoco were identified and characterized by combined biochemical, spectroscopic and structural analyses. This review gives a brief account of the recent progress toward understanding the assembly process of FeMoco, which has identified some important missing pieces of this biosynthetic puzzle.Graphical abstractResearch highlights▶ NifEN accumulates an all-Fe precursor closely resembling the Fe–S core of FeMoco. ▶ NifEN provides a scaffold for the maturation of the all-Fe precursor. ▶ Fe protein inserts Mo and homocitrate in the precursor on NifEN upon ATP hydrolysis. ▶ FeMoco is transferred from NifEN to MoFe protein upon protein–protein interactions. ▶ FeMoco is inserted into MoFe protein via a positively charged insertion funnel.
Co-reporter:Michael A. Blank, Chi Chung Lee, Yilin Hu, Keith O. Hodgson, Britt Hedman, and Markus W. Ribbe
Inorganic Chemistry 2011 Volume 50(Issue 15) pp:7123-7128
Publication Date(Web):June 30, 2011
DOI:10.1021/ic200636k
The iron (Fe) proteins of molybdenum (Mo)-, vanadium (V)-, and iron (Fe)-only nitrogenases are encoded by nifH, vnfH, and anfH, respectively. While the nifH-encoded Fe protein has been extensively studied over recent years, information regarding the properties of the vnfH- and anfH-encoded Fe proteins has remained scarce. Here, we present a combined biochemical, electron paramagnetic resonance (EPR) and X-ray absorption spectroscopy (XAS) analysis of the [Fe4S4] clusters of NifH, VnfH, and AnfH of Azotobacter vinelandii. Our data show that all three Fe proteins contain [Fe4S4] clusters of very similar spectroscopic and geometric structural properties, although NifH differs more from VnfH and AnfH with regard to the electronic structure. These observations have an interesting impact on the theory of the plausible sequence of evolution of nitrogenase Fe proteins. More importantly, the results presented herein provide a platform for future investigations of the differential activities of the three Fe proteins in nitrogenase biosynthesis and catalysis.
Co-reporter:Kresimir Rupnik;Yilin Hu;Aaron W. Fay
JBIC Journal of Biological Inorganic Chemistry 2011 Volume 16( Issue 2) pp:325-332
Publication Date(Web):2011 February
DOI:10.1007/s00775-010-0728-9
NifEN plays a key role in the biosynthesis of the iron–molybdenum cofactor (FeMoco) of nitrogenase. A scaffold protein that hosts the conversion of a FeMoco precursor to a mature cofactor, NifEN can assume three conformations during the process of FeMoco maturation. One, designated ΔnifB NifEN, contains only two permanent [Fe4S4]-like clusters. The second, designated NifENPrecursor, contains the permanent clusters and a precursor form of FeMoco. The third, designated NifEN“FeMoco”, contains the permanent [Fe4S4]-like clusters and a fully complemented, “FeMoco”-like structure. Here, we report a variable-temperature, variable-field magnetic circular dichroism spectroscopic investigation of the electronic structure of the metal clusters in the three forms of dithionite-reduced NifEN. Our data indicate that the permanent [Fe4S4]-like clusters are structurally and electronically conserved in all three NifEN species and exhibit spectral features of classic [Fe4S4]+ clusters; however, they are present in a mixed spin state with a small contribution from the S > ½ spin state. Our results also suggest that both the precursor and “FeMoco” have a conserved Fe/S electronic structure that is similar to the electronic structure of FeMoco in the MoFe protein, and that the “FeMoco” in NifEN“FeMoco” exists, predominantly, in an S = 3/2 spin state with spectral parameters identical to those of FeMoco in the MoFe protein. These observations provide strong support to the outcome of our previous EPR and X-ray absorption spectroscopy/extended X-ray absorption fine structure analysis of the three NifEN species while providing significant new insights into the unique electronic properties of the precursor and “FeMoco” in NifEN.
Co-reporter:Jens T. Kaiser;Yilin Hu;Jared A. Wiig;Douglas C. Rees
Science 2011 Vol 331(6013) pp:91-94
Publication Date(Web):07 Jan 2011
DOI:10.1126/science.1196954
A metalloprotein structure involved in nitrogen fixation offers insight into metal-cluster insertion in nitrogenase.
Co-reporter:Yilin Hu;Chi Chung Lee
Science 2011 Volume 333(Issue 6043) pp:753-755
Publication Date(Web):05 Aug 2011
DOI:10.1126/science.1206883
The molybdenum nitrogenase enzyme can reduce carbon monoxide, albeit inefficiently, in addition to its native substrate, nitrogen.
Co-reporter:Chi Chung Lee;Dr. Yilin Hu;Dr. Markus W. Ribbe
Angewandte Chemie International Edition 2011 Volume 50( Issue 24) pp:5545-5547
Publication Date(Web):
DOI:10.1002/anie.201100869
Co-reporter:Jared A. Wiig;Yilin Hu
PNAS 2011 108 (21 ) pp:8623-8627
Publication Date(Web):2011-05-24
DOI:10.1073/pnas.1102773108
Assembly of nitrogenase FeMoco is one of the key processes in bioinorganic chemistry. NifB and NifEN are two essential elements
immediately adjacent to each other along the biosynthetic pathway of FeMoco. Previously, an 8Fe-precursor of FeMoco was identified
on NifEN; however, the identity of the biosynthetic intermediate on NifB has remained elusive to date. Here, we present a
combined biochemical and spectroscopic investigation of a His-tagged NifEN-B fusion protein of Azotobacter vinelandii. Our data from the EPR and activity analyses confirm the presence of the 8Fe-precursor in the NifEN entity of NifEN-B; whereas
those from the metal, EPR, and UV/Vis experiments reveal the presence of additional [Fe4S4]-type cluster species in the NifB entity of NifEN-B. EPR-, UV/Vis- and metal-based quantitative analyses suggest that the
newly identified cluster species in NifEN-B consist of both SAM-motif (CXXXCXXC)- and non-SAM-motif-bound [Fe4S4]-type clusters. Moreover, EPR and activity experiments indicate that the non-SAM-motif [Fe4S4] cluster is a NifB-bound intermediate of FeMoco assembly, which could be converted to the 8Fe-precursor in a SAM-dependent
mechanism. Combined outcome of this work provides the initial insights into the biosynthetic events of FeMoco on NifB. More
importantly, the full capacity of NifEN-B in FeMoco biosynthesis demonstrates the potential of this fusion protein as an excellent
platform for further investigations of the role of NifB and its interaction with NifEN during the process of FeMoco assembly.
Co-reporter:Aaron W. Fay;Michael A. Blank;Chi Chung Lee;Dr. Yilin Hu; Keith O. Hodgson; Britt Hedman; Markus W. Ribbe
Angewandte Chemie International Edition 2011 Volume 50( Issue 34) pp:7787-7790
Publication Date(Web):
DOI:10.1002/anie.201102724
Co-reporter:Yilin Hu and Markus W. Ribbe
Accounts of Chemical Research 2010 Volume 43(Issue 3) pp:475
Publication Date(Web):December 23, 2009
DOI:10.1021/ar900254x
The (Mo)-nitrogenase is a complex metalloenzyme that catalyzes the key step in the global nitrogen cycle, the reduction of atmospheric dinitrogen (N2) to bioavailable ammonia (NH3), at the iron−molybdenum cofactor (FeMoco) site of its molybdenum−iron (MoFe) protein component. Despite the fundamental significance of biological nitrogen fixation and extensive studies over the past decades, the catalytic mechanism of nitrogenase has not been deciphered. One major challenge for the mechanistic study of nitrogenase is the redox versatility of its FeMoco center. The ability of FeMoco to shuttle between oxidation states in a rapid and unsynchronized manner results in a mixed oxidation state of the cofactor population during turnover. The substrate and the various intermediates can only interact with the FeMoco site in a transient manner, so it is extremely difficult to capture any substrate- or intermediate-bound form of nitrogenase for the direct examination of substrate−enzyme interactions during catalysis. In this Account, we describe the approach of identifying a partially “defective” nitrogenase homologue, one with a slower turnover rate, as a means of overcoming this problem. The NifEN protein complex serves as an ideal candidate for this purpose. It is an α2β2-heterotetramer that contains cluster-binding sites homologous to those found in the MoFe protein: the “P-cluster site” at the interface of the αβ-subunit dimer, which accommodates a [Fe4S4]-type cluster; and the “FeMoco site” within the α-subunit, which houses an all-iron homologue to the FeMoco. Moreover, NifEN mimics the MoFe protein in catalysis: it is capable of reducing acetylene (C2H2) and azide (N3−) in an ATP- and iron (Fe) protein-dependent manner. However, NifEN is unable to reduce proton (H+) and N2, and it is an inefficient enzyme with a restricted electron flux during the turnover. The extremely slow turnover rate of NifEN and the possible “synchronization” of its FeMoco homologue at a certain oxidation level permit the observation of a new S = 1/2 EPR signal upon turnover of C2H2 by NifEN, which is analogous to the signal reported for a MoFe protein variant upon turnover of the same substrate. This result is exciting, because it suggests the possibility of naturally enriching a C2H2-bound form of NifEN for the successful crystallization of the first intermediate-bound nitrogenase homologue. On the other hand, the fact that NifEN represents a partially “defective” homologue of the MoFe protein makes it a promising mutational platform on which a functional MoFe protein equivalent may be reconstructed by introducing the missing features of MoFe protein step-by-step into NifEN. Such a strategy allows us to define the function of each feature and address questions such as the following: What is the function of P-cluster in catalysis? Are Mo and homocitrate the essential constituents of the cofactor in N2 reduction? How does substrate accessibility affect the reactivity of the enzyme? This homologue approach could complement the mechanistic analysis of the nitrogenase MoFe protein, and information derived from both approaches will help achieve the ultimate goal of solving the riddle of biological nitrogen fixation.
Co-reporter:Aaron W. Fay ; Michael A. Blank ; Chi Chung Lee ; Yilin Hu ; Keith O. Hodgson ; Britt Hedman
Journal of the American Chemical Society 2010 Volume 132(Issue 36) pp:12612-12618
Publication Date(Web):August 18, 2010
DOI:10.1021/ja1019657
The cofactors of the Mo- and V-nitrogenases (i.e., FeMoco and FeVco) are homologous metal centers with distinct catalytic properties. So far, there has been only one report on the isolation of FeVco from Azotobacter chroococcum. However, this isolated FeVco species did not carry the full substrate-reducing capacity, as it is unable to restore the N2-reducing ability of the cofactor-deficient MoFe protein. Here, we report the isolation and characterization of a fully active species of FeVco from A. vinelandii. Our metal and activity analyses show that FeVco has been extracted intact, carrying with it the characteristic capacity to reduce C2H2 to C2H6 and, perhaps even more importantly, the ability to reduce N2 to NH3. Moreover, our EPR and XAS/EXAFS investigations indicate that FeVco is similar to, yet distinct from FeMoco in electronic properties and structural topology, which could account for the differences in the reactivity of the two cofactors. The outcome of this study not only permits the proposal of the first EXAFS-based structural model of the isolated FeVco but also lays a foundation for future catalytic and structural investigations of this unique metallocluster.
Co-reporter:Aaron Wolfe Fay, Michael Aaron Blank, Janice Mariko Yoshizawa, Chi Chung Lee, Jared Andrew Wiig, Yilin Hu, Keith Owen Hodgson, Britt Hedman and Markus Walter Ribbe
Dalton Transactions 2010 vol. 39(Issue 12) pp:3124-3130
Publication Date(Web):18 Feb 2010
DOI:10.1039/C000264J
Molybdenum (Mo)-dependent nitrogenase is a complex metalloprotein that catalyzes the biological reduction of dinitrogen (N2) to ammonia (NH3) at the molybdenum-iron cofactor (FeMoco) site of its molybdenum-iron (MoFe) protein component. Here we report the formation of a homocitrate-free, iron-molybdenum (“FeMo”) cluster on the biosynthetic scaffold of FeMoco, NifEN. Such a NifEN-associated “FeMo” cluster exhibits EPR features similar to those of the NifEN-associated, fully-complemented “FeMoco”, which originate from the presence of Mo in both cluster species; however, “FeMo” cluster and “FeMoco” display different temperature-dependent changes in the line shape and the signal intensity of their respective EPR features, which reflect the impact of homocitrate on the redox properties of these clusters. XAS/EXAFS analysis reveals that the Mo centers in both “FeMo” cluster and “FeMoco” are present in a similar coordination environment, although Mo in “FeMo” cluster is more loosely coordinated as compared to that in “FeMoco” with respect to the Mo–O distances in the cluster, likely due to the absence of homocitrate that normally serves as an additional ligand for the Mo in the cluster. Subsequent biochemical investigation of the “FeMo” cluster not only facilitates the determination of the sequence of events in the mobilization of Mo and homocitrate during FeMoco maturation, but also permits the examination of the role of homocitrate in the transfer of FeMoco between NifEN and MoFe protein. Combined outcome of these studies establishes a platform for future structural analysis of the interactions between NifEN and MoFe protein, which will provide useful insights into the mechanism of cluster transfer between the two proteins.
Co-reporter:Yilin Hu, Aaron W. Fay, Chi Chung Lee, Jared A. Wiig and Markus W. Ribbe
Dalton Transactions 2010 vol. 39(Issue 12) pp:2964-2971
Publication Date(Web):11 Jan 2010
DOI:10.1039/B922555B
Nitrogenase catalyzes the nucleotide-dependent conversion of dinitrogen to ammonia at the iron-molybdenum cofactor (FeMoco) center of its molybdenum-iron (MoFe) protein component. Biosynthesis of FeMoco is arguably one of the most complex processes in the field of bioinorganic chemistry, which involves the participation of a number of nif (nitrogen fixing) gene products. One key player in this process, NifEN (encoded by nifE and nifN), is homologous to the MoFe protein with regard to both the primary sequences and the types of the metal centers. Recently, an all-iron precursor has been identified on NifEN, which closely resembles the Fe/S core structure of the mature cofactor. Such a precursor-bound form of NifEN has not only served as an excellent platform for the investigation of FeMoco assembly, but also facilitated the examination of the capacity of NifEN as a catalytic homolog of MoFe protein. This perspective will focus on the recent advances toward elucidating the dual functions of NifEN in nitrogenase assembly and catalysis, and the insights afforded by these advances into the evolution and mechanism of nitrogenase.
Co-reporter:Janice M. Yoshizawa;Aaron W. Fay
JBIC Journal of Biological Inorganic Chemistry 2010 Volume 15( Issue 3) pp:421-428
Publication Date(Web):2010 March
DOI:10.1007/s00775-009-0614-5
The cofactors of Mo-, V-, Fe-dependent nitrogenases are believed to be highly homologous in structure despite the different types of heterometals (Mo, V, and Fe) they contain. Previously, a precursor form of the FeMo cofactor (FeMoco) was captured on NifEN, a scaffold protein for FeMoco biosynthesis. This all-Fe precursor closely resembles the Fe/S core structure of the FeMoco and, therefore, could reasonably serve as a precursor for all nitrogenase cofactors. Here, we report the heterologous incorporation of V and Fe into the NifEN-associated FeMoco precursor. EPR and activity analyses indicate that V and Fe can be inserted at much reduced efficiencies compared with Mo, and incorporation of both V and Fe is enhanced in the presence of homocitrate. Further, native polyacrylamide gel electrophoresis experiments suggest that NifEN undergoes a significant conformational rearrangement upon metal insertion, which allows the subsequent NifEN–MoFe protein interactions and the transfer of the cofactor between the two proteins. The combined outcome of these in vitro studies leads to the proposal of a selective mechanism that is utilized in vivo to maintain the specificity of heterometals in nitrogenase cofactors, which is likely accomplished through the redox regulation of metal mobilization by different Fe proteins (encoded by nifH, vnfH, and anfH, respectively), as well as the differential interactions between these Fe proteins and their respective scaffold proteins (NifEN and VnfEN) in the Mo-, V-, and Fe-dependent nitrogenase systems.
Co-reporter:Yilin Hu;Chi Chung Lee
Science 2010 Volume 329(Issue 5992) pp:642
Publication Date(Web):06 Aug 2010
DOI:10.1126/science.1191455
An enzyme that reduces nitrogen to ammonia can also reduce carbon monoxide to hydrocarbons.
Co-reporter:Janice M. Yoshizawa ; Michael A. Blank ; Aaron W. Fay ; Chi Chung Lee ; Jared A. Wiig ; Yilin Hu ; Keith O. Hodgson ; Britt Hedman
Journal of the American Chemical Society 2009 Volume 131(Issue 26) pp:9321-9325
Publication Date(Web):June 10, 2009
DOI:10.1021/ja9035225
Mo-nitrogenase catalyzes the reduction of dinitrogen to ammonia at the cofactor (i.e., FeMoco) site of its MoFe protein component. Biosynthesis of FeMoco involves NifEN, a scaffold protein that hosts the maturation of a precursor to a mature FeMoco before it is delivered to the target location in the MoFe protein. Previously, we have shown that the NifEN-bound precursor could be converted in vitro to a fully complemented “FeMoco” in the presence of 2 mM dithionite. However, such a conversion was incomplete, and Mo was only loosely associated with the NifEN-bound “FeMoco”. Here we report the optimized maturation of the NifEN-associated precursor in 20 mM dithionite. Activity analyses show that upon the optimal conversion of precursor to “FeMoco”, NifEN is capable of activating a FeMoco-deficient form of MoFe protein to the same extent as the isolated FeMoco. Furthermore, EPR and XAS/EXAFS analyses reveal the presence of a tightly organized Mo site in NifEN-bound “FeMoco”, which allows the observation of a FeMoco-like S = 3/2 EPR signal and the modeling of a NifEN-bound “FeMoco” that adopts a conformation very similar to that of the MoFe protein-associated FeMoco. The sensitivity of FeMoco maturation to dithionite concentration suggests an essential role of redox chemistry in this process, and the optimal potential of dithionite solution could serve as a guideline for future identification of in vivo electron donors for FeMoco maturation.
Co-reporter:Chi Chung Lee;Michael A. Blank;Aaron W. Fay;Janice M. Yoshizawa;Yilin Hu;Keith O. Hodgson;Britt Hedman
PNAS 2009 Volume 106 (Issue 44 ) pp:18474-18478
Publication Date(Web):2009-11-03
DOI:10.1073/pnas.0909149106
The P-cluster of nitrogenase is one of the most complex biological metallocenters known to date. Despite the recent advances
in the chemical synthesis of P-cluster topologs, the biosynthetic mechanism of P-cluster has not been well defined. Here,
we present a combined biochemical, electron paramagnetic resonance, and X-ray absorption spectroscopy/extended X-ray absorption
fine-structure investigation of the maturation process of P-clusters in ΔnifH molybdenum-iron (MoFe) protein. Our data indicate that the previously identified, [Fe4S4]-like cluster pairs in ΔnifH MoFe protein are indeed the precursors to P-clusters, which can be reductively coupled into the mature [Fe8S7] structures in the presence of Fe protein, MgATP, and dithionite. Moreover, our observation of a biphasic maturation pattern
of P-clusters in ΔnifH MoFe protein provides dynamic proof for the previously hypothesized, stepwise assembly mechanism of the two P-clusters in
the α2β2-tetrameric MoFe protein, i.e., one P-cluster is formed in one αβ dimer before the other in the second αβ dimer.
Co-reporter:Yilin Hu;Janice M. Yoshizawa;Aaron W. Fay;Chi Chung Lee;Jared A. Wiig
PNAS 2009 Volume 106 (Issue 40 ) pp:16962-16966
Publication Date(Web):2009-10-06
DOI:10.1073/pnas.0907872106
NifEN is a key player in the biosynthesis of nitrogenase MoFe protein. It not only shares a considerable degree of sequence
homology with the MoFe protein, but also contains clusters that are homologous to those found in the MoFe protein. Here we
present an investigation of the catalytic activities of NifEN. Our data show that NifEN is catalytically competent in acetylene
(C2H2) and azide (N3−) reduction, yet unable to reduce dinitrogen (N2) or evolve hydrogen (H2). Upon turnover, C2H2 gives rise to an additional S = 1/2 signal, whereas N3− perturbs the signal originating from the NifEN-associated FeMoco homolog. Combined biochemical and spectroscopic studies
reveal that N3− can act as either an inhibitor or an activator for the binding and/or reduction of C2H2, while carbon monoxide (CO) is a potent inhibitor for the binding and/or reduction of both N3− and C2H2. Taken together, our results suggest that NifEN is a catalytic homolog of MoFe protein; however, it is only a “skeleton”
version of the MoFe protein, as its associated clusters are simpler in structure and less versatile in function, which, in
turn, may account for its narrower range of substrates and lower activities of substrate reduction. The resemblance of NifEN
to MoFe protein in catalysis points to a plausible, sequential appearance of the two proteins in nitrogenase evolution. More
importantly, the discrepancy between the two systems may provide useful insights into nitrogenase mechanism and allow reconstruction
of a fully functional nitrogenase from the “skeleton” enzyme, NifEN.
Co-reporter:Yilin Hu;Chi Chung Lee
PNAS 2009 Volume 106 (Issue 23 ) pp:9209-9214
Publication Date(Web):2009-06-09
DOI:10.1073/pnas.0904408106
Nitrogenase is an essential metalloenzyme that catalyzes the biological conversion of dinitrogen (N2) to ammonia (NH3). The vanadium (V)-nitrogenase is very similar to the “conventional” molybdenum (Mo)-nitrogenase, yet it holds unique properties
of its own that may provide useful insights into the general mechanism of nitrogenase catalysis. So far, characterization
of the vanadium iron (VFe) protein of Azotobacter vinelandii V-nitrogenase has been focused on 2 incomplete forms of this protein: αβ2 and α2β2, both of which contain the small δ-subunit in minor amounts. Although these studies provided important information about
the V-dependent nitrogenase system, they were hampered by the heterogeneity of the protein samples. Here, we report the isolation
and characterization of a homogeneous, His-tagged form of VFe protein from A. vinelandii. This VFe protein has a previously-unsuspected, α2β2δ4-heterooctameric composition. Further, it contains a P-cluster that is electronically and, perhaps, structurally different
from the P-cluster of molybdenum iron (MoFe) protein. More importantly, it is catalytically distinct from the MoFe protein,
particularly with regard to the mechanism of H2 evolution. A detailed EPR investigation of the origins and interplays of FeV cofactor- and P-cluster-associated signals is
presented herein, which lays the foundation for future kinetic and structural analysis of the VFe protein.
Co-reporter:Yilin Hu, Aaron W. Fay, Chi Chung Lee, Janice Yoshizawa and Markus W. Ribbe
Biochemistry 2008 Volume 47(Issue 13) pp:
Publication Date(Web):March 4, 2008
DOI:10.1021/bi7025003
Assembly of nitrogenase MoFe protein is arguably one of the most complex processes in the field of bioinorganic chemistry, requiring, at least, the participation of nifS, nifU, nifB, nifE, nifN, nifV, nifQ, nifZ, nifH, nifD, and nifK gene products. Previous genetic studies have identified factors involved in MoFe protein assembly; however, the exact functions of these factors and the precise sequence of events during the process have remained unclear until the recent characterization of a number of assembly-related intermediates that provided significant insights into this biosynthetic “black box”. This review summarizes the recent advances in elucidation of the mechanism of FeMoco biosynthesis in four aspects: (1) the ex situ assembly of FeMoco on NifEN, (2) the incorporation of FeMoco into MoFe protein, (3) the in situ assembly of P-cluster on MoFe protein, and (4) the stepwise assembly of MoFe protein.
Co-reporter:Aaron W. Fay, Yilin Hu, Benedikt Schmid, Markus W. Ribbe
Journal of Inorganic Biochemistry 2007 Volume 101(11–12) pp:1630-1641
Publication Date(Web):November 2007
DOI:10.1016/j.jinorgbio.2007.03.013
The final step of FeMo cofactor (FeMoco) assembly involves the insertion of FeMoco into its binding site in the molybdenum–iron (MoFe) protein of nitrogenase. Here we examine the role of His α274 and His α451 of Azotobacter vinelandii MoFe protein in this process. Our results from combined metal, activity, EPR, stability and insertion analyses show that mutations of His α274 and/or His α451, two of the histidines that belong to a so-called His triad, to small uncharged Ala specifically reduce the accumulation of FeMoco in MoFe protein. This observation indicates that the enrichment of histidines at the His triad is important for FeMoco insertion and that the His triad potentially serves as an intermediate docking point for FeMoco through transitory ligand coordination and/or electrostatic interaction.
Co-reporter:Yilin Hu;Aaron W. Fay
JBIC Journal of Biological Inorganic Chemistry 2007 Volume 12( Issue 4) pp:449-460
Publication Date(Web):2007 May
DOI:10.1007/s00775-006-0199-1
The assembly of the complex iron–molybdenum cofactor (FeMoco) of nitrogenase molybdenum–iron (MoFe) protein has served as one of the central topics in the field of bioinorganic chemistry for decades. Here we examine the role of a MoFe protein residue (His α362) in FeMoco insertion, the final step of FeMoco biosynthesis where FeMoco is incorporated into its binding site in the MoFe protein. Our data from combined metal, activity and electron paramagnetic resonance analyses show that mutations of His α362 to small uncharged Ala or negatively charged Asp result in significantly reduced FeMoco accumulation in MoFe protein, indicating that His α362 plays a key role in the process of FeMoco insertion. Given the strategic location of His α362 at the entry point of the FeMoco insertion funnel, this residue may serve as one of the initial docking points for FeMoco insertion through transient ligand coordination and/or electrostatic interaction.
Co-reporter:Markus W. Ribbe;Yilin Hu;Aaron W. Fay;Chi Chung Lee
PNAS 2007 Volume 104 (Issue 25 ) pp:10424-10429
Publication Date(Web):2007-06-19
DOI:10.1073/pnas.0704297104
Biosynthesis of nitrogenase P-cluster has attracted considerable attention because it is biologically important and chemically
unprecedented. Previous studies suggest that P-cluster is formed from a precursor consisting of paired [4Fe–4S]-like clusters
and that P-cluster is assembled stepwise on MoFe protein, i.e., one cluster is assembled before the other. Here, we specifically
tackle the assembly of the second P-cluster by combined biochemical and spectroscopic approaches. By using a P-cluster maturation
assay that is based on purified components, we show that the maturation of the second P-cluster requires the concerted action
of NifZ, Fe protein, and MgATP and that the action of NifZ is required before that of Fe protein/MgATP, suggesting that NifZ
may act as a chaperone that facilitates the subsequent action of Fe protein/MgATP. Furthermore, we provide spectroscopic evidence
that the [4Fe–4S] cluster-like fragments can be converted to P-clusters, thereby firmly establishing the physiological relevance
of the previously identified P-cluster precursor.
Co-reporter:Yilin Hu;Mary C. Corbett;Jerome A. Webber;Keith O. Hodgson;Aaron W. Fay;Britt Hedman
PNAS 2006 Volume 103 (Issue 46 ) pp:17125-17130
Publication Date(Web):2006-11-14
DOI:10.1073/pnas.0602651103
The Fe protein is indispensable for nitrogenase catalysis and biosynthesis. However, its function in iron-molybdenum cofactor
(FeMoco) biosynthesis has not been clearly defined. Here we show that the Fe protein can act as a Mo/homocitrate insertase
that mobilizes Mo/homocitrate for the maturation of FeMoco precursor on NifEN. Further, we establish that Mo/homocitrate mobilization
by the Fe protein likely involves hydrolysis of MgATP and protein–protein interaction between the Fe protein and NifEN. Our
findings not only clarify the role of the Fe protein in FeMoco assembly and assign another function to this multitask enzyme
but also provide useful insights into a mechanism of metal trafficking required for the assembly of complex metalloproteins
such as nitrogenase.
Co-reporter:Yilin Hu;Aaron W. Fay;Mary C. Corbett;Jerome A. Webber;Keith O. Hodgson;Britt Hedman
PNAS 2006 Volume 103 (Issue 46 ) pp:17119-17124
Publication Date(Web):2006-11-14
DOI:10.1073/pnas.0602647103
FeMo cofactor (FeMoco) biosynthesis is one of the most complicated processes in metalloprotein biochemistry. Here we show
that Mo and homocitrate are incorporated into the Fe/S core of the FeMoco precursor while it is bound to NifEN and that the
resulting fully complemented, FeMoco-like cluster is transformed into a mature FeMoco upon transfer from NifEN to MoFe protein
through direct protein–protein interaction. Our findings not only clarify the process of FeMoco maturation, but also provide
useful insights into the other facets of nitrogenase chemistry.
Co-reporter:Mary C. Corbett;Yilin Hu;Aaron W. Fay;Britt Hedman;Keith O. Hodgson
PNAS 2006 Volume 103 (Issue 5 ) pp:1238-1243
Publication Date(Web):2006-01-31
DOI:10.1073/pnas.0507853103
The iron-molybdenum cofactor (FeMoco) of the nitrogenase MoFe protein is a highly complex metallocluster that provides the
catalytically essential site for biological nitrogen fixation. FeMoco is assembled outside the MoFe protein in a stepwise
process requiring several components, including NifB-co, an iron- and sulfur-containing FeMoco precursor, and NifEN, an intermediary
assembly protein on which NifB-co is presumably converted to FeMoco. Through the comparison of Azotobacter vinelandii strains expressing the NifEN protein in the presence or absence of the nifB gene, the structure of a NifEN-bound FeMoco precursor has been analyzed by x-ray absorption spectroscopy. The results provide
physical evidence to support a mechanism for FeMoco biosynthesis. The NifEN-bound precursor is found to be a molybdenum-free
analog of FeMoco and not one of the more commonly suggested cluster types based on a standard [4Fe–4S] architecture. A facile
scheme by which FeMoco and alternative, non-molybdenum-containing nitrogenase cofactors are constructed from this common precursor
is presented that has important implications for the biosynthesis and biomimetic chemical synthesis of FeMoco.
Co-reporter:Yilin Hu;Mary C. Corbett;Aaron W. Fay;Jerome A. Webber;Britt Hedman;Keith O. Hodgson
PNAS 2005 Volume 102 (Issue 39 ) pp:13825-13830
Publication Date(Web):2005-09-27
DOI:10.1073/pnas.0506967102
Nitrogenase is a multicomponent metalloenzyme that catalyzes the conversion of atmospheric dinitrogen to ammonia. For decades,
it has been generally believed that the [8Fe-7S] P-cluster of nitrogenase component 1 is indispensable for nitrogenase activity.
In this study, we identified two catalytically active P-cluster variants by activity assays, metal analysis, and EPR spectroscopic
studies. Further, we showed that both P-cluster variants resemble [4Fe-4S]-like centers based on x-ray absorption spectroscopic
experiments. We believe that our findings challenge the dogma that the standard P-cluster is the only cluster species capable
of supporting substrate reduction at the FeMo cofactor and provide important insights into the general mechanism of nitrogenase
catalysis and assembly.
Co-reporter:Yilin Hu;Aaron W. Fay;
Proceedings of the National Academy of Sciences 2005 102(9) pp:3236-3241
Publication Date(Web):February 22, 2005
DOI:10.1073/pnas.0409201102
The biosynthesis of the FeMo cofactor (FeMoco) of Azotobacter vinelandii nitrogenase presumably starts with the production of its Fe/S core by NifB (the nifB gene product). This core is subsequently processed on the α2β2 tetrameric NifEN complex (formed by the nifE and nifN gene products). In this article, we identify a NifEN-bound FeMoco precursor form that can be converted to fully assembled
FeMoco in a so-called FeMoco-maturation assay containing only purified components. We also establish that only molybdate,
homocitrate, MgATP, and Fe protein are essential for FeMoco maturation. The FeMoco-maturation assay described here will further
address the remaining questions related to the assembly mechanism of the ever-intriguing FeMoco.
Co-reporter:Aaron W. Fay, Yilin Hu, Benedikt Schmid, Markus W. Ribbe
Journal of Inorganic Biochemistry (November 2007) Volume 101(11–12) pp:1630-1641
Publication Date(Web):1 November 2007
DOI:10.1016/j.jinorgbio.2007.03.013
The final step of FeMo cofactor (FeMoco) assembly involves the insertion of FeMoco into its binding site in the molybdenum–iron (MoFe) protein of nitrogenase. Here we examine the role of His α274 and His α451 of Azotobacter vinelandii MoFe protein in this process. Our results from combined metal, activity, EPR, stability and insertion analyses show that mutations of His α274 and/or His α451, two of the histidines that belong to a so-called His triad, to small uncharged Ala specifically reduce the accumulation of FeMoco in MoFe protein. This observation indicates that the enrichment of histidines at the His triad is important for FeMoco insertion and that the His triad potentially serves as an intermediate docking point for FeMoco through transitory ligand coordination and/or electrostatic interaction.
Co-reporter:Yilin Hu, Markus W. Ribbe
Biochimica et Biophysica Acta (BBA) - Bioenergetics (August–September 2013) Volume 1827(Issues 8–9) pp:1112-1122
Publication Date(Web):August–September 2013
DOI:10.1016/j.bbabio.2012.12.001
Co-reporter:Kresimir Rupnik ; Chi Chung Lee ; Yilin Hu ; Markus W. Ribbe ;Brian J. Hales
Journal of the American Chemical Society () pp:
Publication Date(Web):April 13, 2011
DOI:10.1021/ja201384w
Two proteins involved in nitrogen fixation contain ferredoxin-type [4Fe4S] clusters that exist in paramagnetic ground state upon oxidation, a property never observed since the discovery of ferredoxins 50 years ago. This unique characteristic suggests a specific coupling in these clusters necessary for nitrogen fixation and implies an evolutionary connection between the clusters in the two proteins.
Co-reporter:Aaron Wolfe Fay, Michael Aaron Blank, Janice Mariko Yoshizawa, Chi Chung Lee, Jared Andrew Wiig, Yilin Hu, Keith Owen Hodgson, Britt Hedman and Markus Walter Ribbe
Dalton Transactions 2010 - vol. 39(Issue 12) pp:NaN3130-3130
Publication Date(Web):2010/02/18
DOI:10.1039/C000264J
Molybdenum (Mo)-dependent nitrogenase is a complex metalloprotein that catalyzes the biological reduction of dinitrogen (N2) to ammonia (NH3) at the molybdenum-iron cofactor (FeMoco) site of its molybdenum-iron (MoFe) protein component. Here we report the formation of a homocitrate-free, iron-molybdenum (“FeMo”) cluster on the biosynthetic scaffold of FeMoco, NifEN. Such a NifEN-associated “FeMo” cluster exhibits EPR features similar to those of the NifEN-associated, fully-complemented “FeMoco”, which originate from the presence of Mo in both cluster species; however, “FeMo” cluster and “FeMoco” display different temperature-dependent changes in the line shape and the signal intensity of their respective EPR features, which reflect the impact of homocitrate on the redox properties of these clusters. XAS/EXAFS analysis reveals that the Mo centers in both “FeMo” cluster and “FeMoco” are present in a similar coordination environment, although Mo in “FeMo” cluster is more loosely coordinated as compared to that in “FeMoco” with respect to the Mo–O distances in the cluster, likely due to the absence of homocitrate that normally serves as an additional ligand for the Mo in the cluster. Subsequent biochemical investigation of the “FeMo” cluster not only facilitates the determination of the sequence of events in the mobilization of Mo and homocitrate during FeMoco maturation, but also permits the examination of the role of homocitrate in the transfer of FeMoco between NifEN and MoFe protein. Combined outcome of these studies establishes a platform for future structural analysis of the interactions between NifEN and MoFe protein, which will provide useful insights into the mechanism of cluster transfer between the two proteins.
Co-reporter:Yilin Hu, Aaron W. Fay, Chi Chung Lee, Jared A. Wiig and Markus W. Ribbe
Dalton Transactions 2010 - vol. 39(Issue 12) pp:NaN2971-2971
Publication Date(Web):2010/01/11
DOI:10.1039/B922555B
Nitrogenase catalyzes the nucleotide-dependent conversion of dinitrogen to ammonia at the iron-molybdenum cofactor (FeMoco) center of its molybdenum-iron (MoFe) protein component. Biosynthesis of FeMoco is arguably one of the most complex processes in the field of bioinorganic chemistry, which involves the participation of a number of nif (nitrogen fixing) gene products. One key player in this process, NifEN (encoded by nifE and nifN), is homologous to the MoFe protein with regard to both the primary sequences and the types of the metal centers. Recently, an all-iron precursor has been identified on NifEN, which closely resembles the Fe/S core structure of the mature cofactor. Such a precursor-bound form of NifEN has not only served as an excellent platform for the investigation of FeMoco assembly, but also facilitated the examination of the capacity of NifEN as a catalytic homolog of MoFe protein. This perspective will focus on the recent advances toward elucidating the dual functions of NifEN in nitrogenase assembly and catalysis, and the insights afforded by these advances into the evolution and mechanism of nitrogenase.
Co-reporter:Yilin Hu, Chi Chung Lee and Markus W. Ribbe
Dalton Transactions 2012 - vol. 41(Issue 4) pp:NaN1127-1127
Publication Date(Web):2011/11/18
DOI:10.1039/C1DT11535A
Nitrogenase catalyzes the biological conversion of atmospheric dinitrogen to bioavailable ammonia. The molybdenum (Mo)- and vanadium (V)-dependent nitrogenases are two homologous members of this metalloenzyme family. However, despite their similarities in structure and function, the characterization of V-nitrogenase has taken a much longer and more winding path than that of its Mo-counterpart. From the initial discovery of this nitrogen-fixing system, to the recent finding of its CO-reducing capacity, V-nitrogenase has proven to be a two-hit wonder in the over-a-century-long research of nitrogen fixation. This perspective provides a brief account of the catalytic function and structural basis of V-nitrogenase, as well as a short discussion of the theoretical and practical potentials of this unique metalloenzyme.
.jpg)
![[5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl [hydroxy-[(3,4,5-trihydroxyoxolan-2-yl)methoxy]phosphoryl] Hydrogen Phosphate](http://img.cochemist.com/ccimg/20800/20762-30-5.png)
![[5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl [hydroxy-[(3,4,5-trihydroxyoxolan-2-yl)methoxy]phosphoryl] Hydrogen Phosphate](http://img.cochemist.com/ccimg/20800/20762-30-5_b.png)