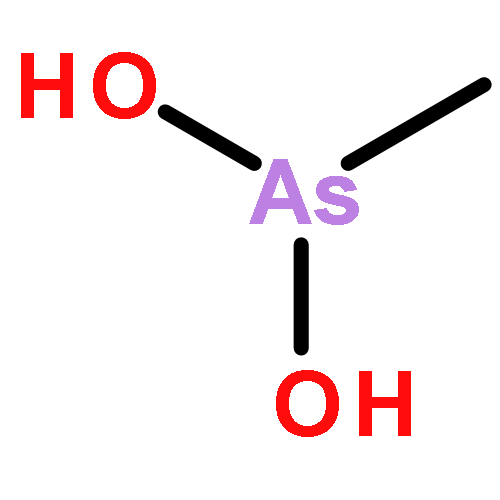
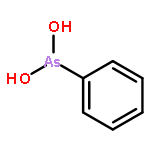
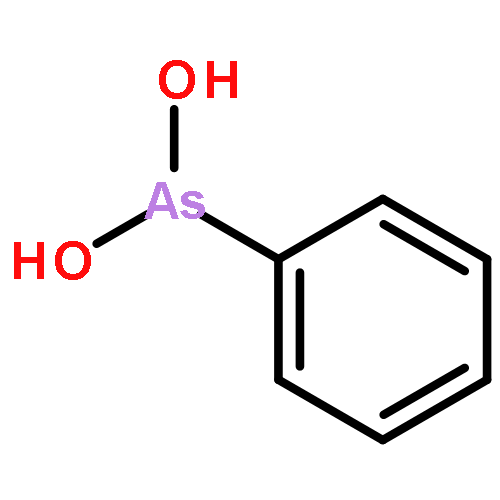
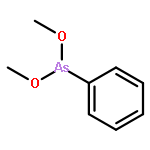
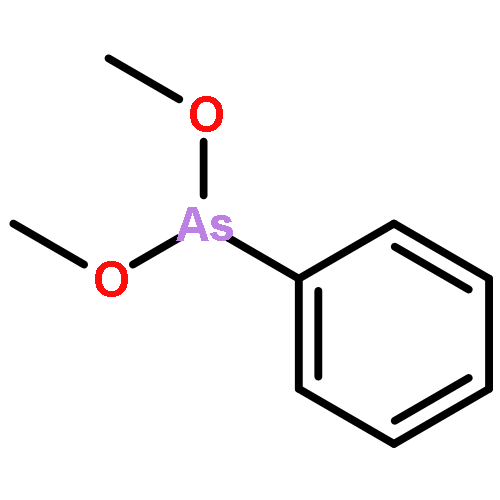
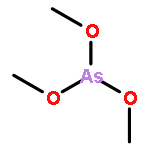
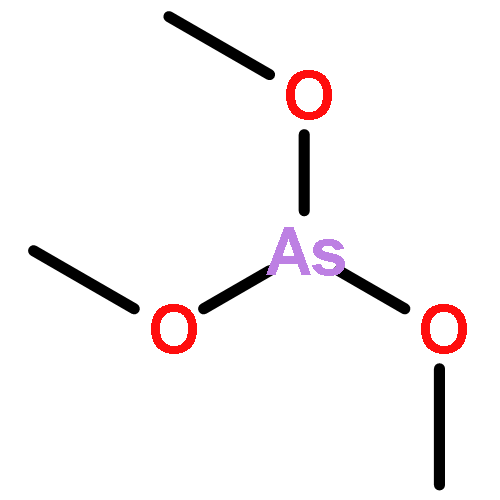
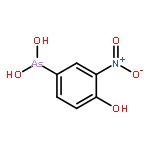
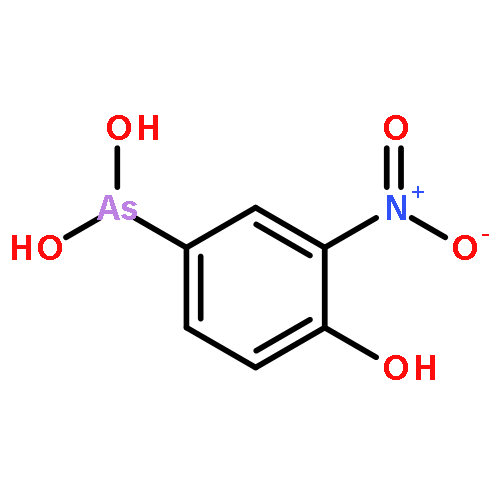
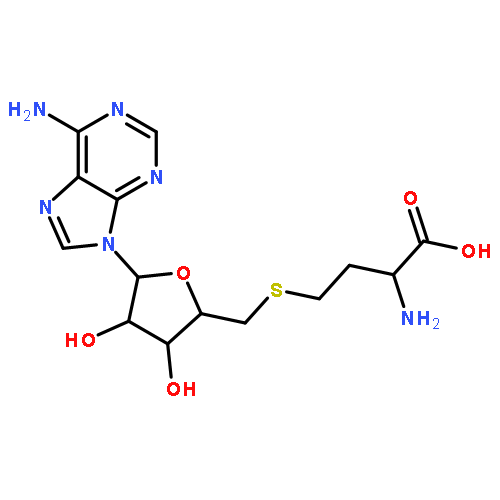
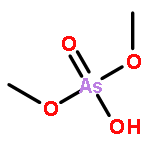
Co-reporter:Jiaojiao Li, Charles Packianathan, Toby G. Rossman, and Barry P. Rosen
Chemical Research in Toxicology July 17, 2017 Volume 30(Issue 7) pp:1481-1481
Publication Date(Web):May 24, 2017
DOI:10.1021/acs.chemrestox.7b00113
Arsenic methylation, the primary biotransformation in the human body, is catalyzed by the enzyme As(III) S-adenosylmethionine (SAM) methyltransferases (hAS3MT). This process is thought to be protective from acute high-level arsenic exposure. However, with long-term low-level exposure, hAS3MT produces intracellular methylarsenite (MAs(III)) and dimethylarsenite (DMAs(III)), which are considerably more toxic than inorganic As(III) and may contribute to arsenic-related diseases. Several single nucleotide polymorphisms (SNPs) in putative regulatory elements of the hAS3MT gene have been shown to be protective. In contrast, three previously identified exonic SNPs (R173W, M287T, and T306I) may be deleterious. The goal of this study was to examine the effect of single amino acid substitutions in hAS3MT on the activity of the enzyme that might explain their contributions to adverse health effects of environmental arsenic. We identified five additional intragenic variants in hAS3MT (H51R, C61W, I136T, W203C, and R251H). We purified the eight polymorphic hAS3MT proteins and characterized their enzymatic properties. Each enzyme had low methylation activity through decreased affinity for substrate, lower overall rates of catalysis, or lower stability. We propose that amino acid substitutions in hAS3MT with decreased catalytic activity lead to detrimental responses to environmental arsenic and may increase the risk of arsenic-related diseases.
Co-reporter:Jian Chen and Barry P. Rosen
Environmental Science & Technology 2016 Volume 50(Issue 15) pp:7956-7963
Publication Date(Web):July 1, 2016
DOI:10.1021/acs.est.6b00235
Microbes play a critical role in the global arsenic biogeocycle. Most studies have focused on redox cycling of inorganic arsenic in bacteria and archaea. The parallel cycles of organoarsenical biotransformations are less well characterized. Here we describe organoarsenical biotransformations in the environmental microbe Shewanella putrefaciens. Under aerobic growth conditions, S. putrefaciens reduced the herbicide MSMA (methylarsenate or MAs(V)) to methylarsenite (MAs(III)). Even though it does not contain an arsI gene, which encodes the ArsI C–As lyase, S. putrefaciens demethylated MAs(III) to As(III). It cleaved the C–As bond in aromatic arsenicals such as the trivalent forms of the antimicrobial agents roxarsone (Rox(III)), nitarsone (Nit(III)) and phenylarsenite (PhAs(III)), which have been used as growth promoters for poultry and swine. S. putrefaciens thiolated methylated arsenicals, converting MAs(V) into the more toxic metabolite monomethyl monothioarsenate (MMMTAs(V)), and transformed dimethylarsenate (DMAs(V)) into dimethylmonothioarsenate (DMMTAs(V)). It also reduced the nitro groups of Nit(V), forming p-aminophenyl arsenate (p-arsanilic acid or p-AsA(V)), and Rox(III), forming 3-amino-4-hydroxybenzylarsonate (3A4HBzAs(V)). Elucidation of organoarsenical biotransformations by S. putrefaciens provides a holistic appreciation of how these environmental pollutants are degraded.
Co-reporter:Hui Dong, Mahendra Madegowda, Adel Nefzi, Richard A. Houghten, Marc A. Giulianotti, and Barry P. Rosen
Chemical Research in Toxicology 2015 Volume 28(Issue 12) pp:2419
Publication Date(Web):November 17, 2015
DOI:10.1021/acs.chemrestox.5b00432
Arsenic is the most ubiquitous environmental toxin and carcinogen. Long-term exposure to arsenic is associated with human diseases including cancer, cardiovascular disease, and diabetes. Human As(III) S-adenosylmethionine (SAM) methyltransferases (hAS3MT) methylates As(III) to trivalent mono- and dimethyl species that are more toxic and potentially more carcinogenic than inorganic arsenic. Modulators of hAS3MT activity may be useful for the prevention or treatment of arsenic-related diseases. Using a newly developed high-throughput assay for hAS3MT activity, we identified 10 novel noncompetitive small molecule inhibitors. In silico docking analysis with the crystal structure of an AS3MT orthologue suggests that the inhibitors bind in a cleft between domains that is distant from either the As(III) or SAM binding sites. This suggests the presence of a possible allosteric and regulatory site in the enzyme. These inhibitors may be useful tools for future research in arsenic metabolism and are the starting-point for the development of drugs against hAS3MT.
Co-reporter:Hui Dong, Wenzhong Xu, Jitesh K. Pillai, Charles Packianathan, Barry P. Rosen
Analytical Biochemistry 2015 480() pp: 67-73
Publication Date(Web):1 July 2015
DOI:10.1016/j.ab.2015.04.011
Arsenic is a naturally existing toxin and carcinogen. As(III) S-adenosylmethionine methyltransferases (AS3MT in mammals and ArsM in microbes) methylate As(III) three times in consecutive steps and play a central role in arsenic metabolism from bacteria to humans. Current assays for arsenic methylation are slow, laborious, and expensive. Here we report the development of two in vitro assays for AS3MT activity that are rapid, sensitive, convenient, and relatively inexpensive and can be adapted for high-throughput assays. The first assay measures As(III) binding by the quenching of the protein fluorescence of a single-tryptophan derivative of an AS3MT ortholog. The second assay utilizes time-resolved fluorescence resonance energy transfer to directly measure the conversion of the AS3MT substrate, S-adenosylmethionine, to S-adenosylhomocysteine catalyzed by AS3MT. These two assays are complementary, one measuring substrate binding and the other catalysis, making them useful tools for functional studies and future development of drugs to prevent arsenic-related diseases.
Co-reporter:Jian Chen, Guo-Xin Sun, Xiao-Xue Wang, Víctor de Lorenzo, Barry P. Rosen, and Yong-Guan Zhu
Environmental Science & Technology 2014 Volume 48(Issue 17) pp:10337-10344
Publication Date(Web):August 5, 2014
DOI:10.1021/es502230b
Even though arsenic is one of the most widespread environmental carcinogens, methods of remediation are still limited. In this report we demonstrate that a strain of Pseudomonas putida KT2440 endowed with chromosomal expression of the arsM gene encoding the As(III) S-adenosylmethionine (SAM) methyltransfase from Rhodopseudomonas palustris to remove arsenic from contaminated soil. We genetically engineered the P. putida KT2440 with stable expression of an arsM-gfp fusion gene (GE P. putida), which was inserted into the bacterial chromosome. GE P. putida showed high arsenic methylation and volatilization activity. When exposed to 25 μM arsenite or arsenate overnight, most inorganic arsenic was methylated to the less toxic methylated arsenicals methylarsenate (MAs(V)), dimethylarsenate (DMAs(V)) and trimethylarsine oxide (TMAs(V)O). Of total added arsenic, the species were about 62 ± 2.2% DMAs(V), 25 ± 1.4% MAs(V) and 10 ± 1.2% TMAs(V)O. Volatilized arsenicals were trapped, and the predominant species were dimethylarsine (Me2AsH) (21 ± 1.0%) and trimethylarsine (TMAs(III)) (10 ± 1.2%). At later times, more DMAs(V) and volatile species were produced. Volatilization of Me2AsH and TMAs(III) from contaminated soil is thus possible with this genetically engineered bacterium and could be instrumental as an agent for reducing the inorganic arsenic content of soil and agricultural products.
Co-reporter:Jian Chen, Samio Sun, Chen-Zhong Li, Yong-Guan Zhu, and Barry P. Rosen
Environmental Science & Technology 2014 Volume 48(Issue 2) pp:1141-1147
Publication Date(Web):December 20, 2013
DOI:10.1021/es4038319
The toxic metalloid arsenic is widely distributed in food, water, and soil. While inorganic arsenic enters the environment primarily from geochemical sources, methylarsenicals either result from microbial biotransformation of inorganic arsenic or are introduced anthropogenically. Methylarsenicals such as monosodium methylarsonic acid (MSMA) have been extensively utilized as herbicides, and aromatic arsenicals such as roxarsone (Rox) are used as growth promoters for poultry and swine. Organoarsenicals are degraded to inorganic arsenic. The toxicological effects of arsenicals depend on their oxidation state, chemical composition, and bioavailability. Here we report that the active forms are the trivalent arsenic-containing species. We constructed a whole-cell biosensor utilizing a modified ArsR repressor that is highly selective toward trivalent methyl and aromatic arsenicals, with essentially no response to inorganic arsenic. The biosensor was adapted for in vitro detection of organoarsenicals using fluorescence anisotropy of ArsR–DNA interactions. It detects bacterial biomethylation of inorganic arsenite both in vivo and in vitro with detection limits of 10–7 M and linearity to 10–6 M for phenylarsenite and 5 × 10–6 M for methylarsenite. The biosensor detects reduced forms of MSMA and roxarsone and offers a practical, low cost method for detecting activate forms and breakdown products of organoarsenical herbicides and growth promoters.
Co-reporter:Dharmendra S. Dheeman, Charles Packianathana, Jitesh K. Pillai, and Barry P. Rosen
Chemical Research in Toxicology 2014 Volume 27(Issue 12) pp:2012
Publication Date(Web):November 10, 2014
DOI:10.1021/tx500439e
Co-reporter:Dharmendra S. Dheeman, Charles Packianathan, Jitesh K. Pillai, and Barry P. Rosen
Chemical Research in Toxicology 2014 Volume 27(Issue 11) pp:1979
Publication Date(Web):October 17, 2014
DOI:10.1021/tx500313k
A synthetic gene encoding human As(III) S-adenosylmethionine (SAM) methyltransferase (hAS3MT) was expressed, and the purified enzyme was characterized. The synthetic enzyme is considerably more active than a cDNA-expressed enzyme using endogenous reductants thioredoxin (Trx), thioredoxin reductase (TR), NADPH, and reduced glutathione (GSH). Each of the seven cysteines (the four conserved residues, Cys32, Cys61, Cys156, and Cys206, and nonconserved, Cys72, Cys85, and Cys250) was individually changed to serine. The nonconserved cysteine derivates were still active. None of the individual C32S, C61S, C156S, and C206S derivates were able to methylate As(III). However, the C32S and C61S enzymes retained the ability to methylate MAs(III). These observations suggest that Cys156 and Cys206 play a different role in catalysis than that of Cys32 and Cys61. A homology model built on the structure of a thermophilic orthologue indicates that Cys156 and Cys206 form the As(III) binding site, whereas Cys32 and Cys61 form a disulfide bond. Two observations shed light on the pathway of methylation. First, binding assays using the fluorescence of a single-tryptophan derivative indicate that As(GS)3 binds to the enzyme much faster than inorganic As(III). Second, the major product of the first round of methylation is MAs(III), not MAs(V), and remains enzyme-bound until it is methylated a second time. We propose a new pathway for hAS3MT catalysis that reconciles the hypothesis of Challenger ((1947) Sci. Prog., 35, 396–416) with the pathway proposed by Hayakawa et al. ((2005) Arch. Toxicol., 79, 183–191). The products are the more toxic and more carcinogenic trivalent methylarsenicals, but arsenic undergoes oxidation and reduction as enzyme-bound intermediates.
Co-reporter:S. Venkadesh Nadar;Masafumi Yoshinaga;Palani Kavelu;Banumathi Sankaran
Acta Crystallographica Section F 2014 Volume 70( Issue 6) pp:761-764
Publication Date(Web):
DOI:10.1107/S2053230X14008814
Arsenic is a ubiquitous and carcinogenic environmental element that enters the biosphere primarily from geochemical sources, but also through anthropogenic activities. Microorganisms play an important role in the arsenic biogeochemical cycle by biotransformation of inorganic arsenic into organic arsenicals and vice versa. ArsI is a microbial nonheme ferrous-dependent dioxygenase that transforms toxic methylarsonous acid to the less toxic inorganic arsenite by C–As bond cleavage. An ArsI ortholog from the thermophilic bacterium Thermomonospora curvata was expressed, purified and crystallized. The crystals diffracted to 1.46 Å resolution and belonged to space group P43212 or its enantiomer P41212, with unit-cell parameters a = b = 42.2, c = 118.5 Å.
Co-reporter:Charles Packianathan;Jitesh K. Pillai;Ahmed Riaz;Palani Kavelu;Banumathi Sankaran
Acta Crystallographica Section F 2014 Volume 70( Issue 10) pp:1385-1388
Publication Date(Web):
DOI:10.1107/S2053230X14018469
Arsenic is one the most toxic environmental substances. Arsenic is ubiquitous in water, soil and food, and ranks first on the Environmental Protection Agency's Superfund Priority List of Hazardous Substances. Arsenic(III) S-adenosylmethionine methyltransferases (AS3MT in animals and ArsM in microbes) are key enzymes of arsenic biotransformation, catalyzing the methylation of inorganic arsenite to give methyl, dimethyl and trimethyl products. Arsenic methyltransferases are found in members of every kingdom from bacteria to humans (EC 2.1.1.137). In the human liver, hAS3MT converts inorganic arsenic into more toxic and carcinogenic forms. CrArsM, an ortholog of hAS3MT from the eukaryotic green alga Chlamydomonas reinhardtii, was purified by chemically synthesizing the gene and expressing it in Escherichia coli. Synthetic purified CrArsM was crystallized in an unliganded form. Crystals were obtained by the hanging-drop vapor-diffusion method. The crystals belonged to space group R3:H, with unit-cell parameters a = b = 157.8, c = 95.4 Å, γ = 120° and two molecules in the asymmetric unit. Complete data sets were collected and processed to a resolution of 2.40 Å.
Co-reporter:Kavitha Marapakala, Jie Qin, and Barry P. Rosen
Biochemistry 2012 Volume 51(Issue 5) pp:
Publication Date(Web):January 17, 2012
DOI:10.1021/bi201500c
The enzyme As(III) S-adenosylmethionine methyltransferase (EC 2.1.1.137) (ArsM or AS3MT) is found in members of every kingdom, from bacteria to humans. In these enzymes, there are three conserved cysteine residues at positions 72, 174, and 224 in the CmArsM orthologue from the thermophilic eukaryotic alga Cyanidioschyzon sp. 5508. Substitution of any of the three led to loss of As(III) methylation. In contrast, a C72A mutant still methylated trivalent methylarsenite [MAs(III)]. Protein fluorescence of a single-tryptophan mutant reported binding of As(III) or MAs(III). As(GS)3 and MAs(GS)2 bound significantly faster than As(III), suggesting that the glutathionylated arsenicals are preferred substrates for the enzyme. Protein fluorescence also reported binding of Sb(III), and the purified enzyme methylated and volatilized Sb(III). The results suggest that all three cysteine residues are necessary for the first step in the reaction, As(III) methylation, but that only Cys174 and Cys224 are required for the second step, methylation of MAs(III) to dimethylarsenite [DMAs(III)]. The rate-limiting step was identified as the conversion of DMAs(III) to trimethylarsine, and DMAs(III) accumulates as the principal product.
Co-reporter:A. Abdul Ajees, Kavitha Marapakala, Charles Packianathan, Banumathi Sankaran, and Barry P. Rosen
Biochemistry 2012 Volume 51(Issue 27) pp:
Publication Date(Web):June 19, 2012
DOI:10.1021/bi3004632
Enzymatic methylation of arsenic is a detoxification process in microorganisms but in humans may activate the metalloid to more carcinogenic species. We describe the first structure of an As(III) S-adenosylmethionine methyltransferase by X-ray crystallography that reveals a novel As(III) binding domain. The structure of the methyltransferase from the thermophilic eukaryotic alga Cyanidioschyzon merolae reveals the relationship between the arsenic and S-adenosylmethionine binding sites to a final resolution of ∼1.6 Å. As(III) binding causes little change in conformation, but binding of SAM reorients helix α4 and a loop (residues 49–80) toward the As(III) binding domain, positioning the methyl group for transfer to the metalloid. There is no evidence of a reductase domain. These results are consistent with previous suggestions that arsenic remains trivalent during the catalytic cycle. A homology model of human As(III) S-adenosylmethionine methyltransferase with the location of known polymorphisms was constructed. The structure provides insights into the mechanism of substrate binding and catalysis.
Co-reporter:A. Abdul Ajees;Jianbo Yang
BioMetals 2011 Volume 24( Issue 3) pp:391-399
Publication Date(Web):2011 June
DOI:10.1007/s10534-010-9398-x
Arsenic, a toxic metalloid widely existing in the environment, causes a variety of health problems. The ars operon encoded by Escherichia coli plasmid R773 has arsD and arsA genes, where ArsA is an ATPase that is the catalytic subunit of the ArsAB As(III) extrusion pump, and ArsD is an arsenic chaperone for ArsA. ArsD transfers As(III) to ArsA and increases the affinity of ArsA for As(III), allowing resistance to environmental concentrations of arsenic. Cys12, Cys13 and Cys18 in ArsD form a three sulfur-coordinated As(III) binding site that is essential for metallochaperone activity. ATP hydrolysis by ArsA is required for transfer of As(III) from ArsD to ArsA, suggesting that transfer occurs with a conformation of ArsA that transiently forms during the catalytic cycle. The 1.4 Å x-ray crystal structure of ArsD shows a core of four β-strands flanked by four α-helices in a thioredoxin fold. Docking of ArsD with ArsA was modeled in silico. Independently ArsD mutants exhibiting either weaker or stronger interaction with ArsA were selected. The locations of the mutations mapped on the surface of ArsD are consistent with the docking model. The results suggest that the interface with ArsA involves one surface of α1 helix and metalloid binding site of ArsD.
Co-reporter:Jianbo Yang, Swati Rawat, Timothy L. Stemmler and Barry P. Rosen
Biochemistry 2010 Volume 49(Issue 17) pp:
Publication Date(Web):April 2, 2010
DOI:10.1021/bi100026a
ArsD is a metallochaperone that delivers trivalent metalloids [As(III) or Sb(III)] to the ArsA ATPase, the catalytic subunit of the ArsAB pump encoded by the arsRDABC operon of Escherichia coli plasmid R773. Interaction with ArsD increases the affinity of ArsA for As(III), conferring resistance to environmental concentrations of arsenic. Previous genetic analysis suggested that ArsD residues Cys12, Cys13, and Cys18 are involved in the transfer of As(III) to ArsA. Here X-ray absorption spectroscopy was used to show that As(III) is coordinated with three sulfur atoms, consistent with the three cysteine residues forming the As(III) binding site. Two single-tryptophan derivatives of ArsD exhibited quenching of intrinsic protein fluorescence upon binding of As(III) or Sb(III), which allowed estimation of the rates of binding and affinities for metalloids. Substitution of Cys12, Cys13, or Cys18 decreased the affinity for As(III) more than 10-fold. Reduced glutathione greatly increased the rate of binding of As(III) to ArsD but did not affect binding of As(III) to ArsA. This suggests that in vivo cytosolic As(III) might be initially bound to GSH and transferred to ArsD and then to ArsAB, which pumps the metalloid out of the cell. The As(III) chelator dimercaptosuccinic acid did not block the transfer from ArsD to ArsA, consistent with channeling of the metalloid from one protein to the other, as opposed to release and rebinding of the metalloid. Finally, transfer of As(III) from ArsD to ArsA occurred in the presence of MgATP at 23 °C but not at 4 °C. Neither MgADP nor MgATP-γ-S could replace MgATP. These results suggest that transfer occurs with a conformation of ArsA that transiently forms during the catalytic cycle.
Co-reporter:Jun Ye, A. Abdul Ajees, Jianbo Yang and Barry P. Rosen
Biochemistry 2010 Volume 49(Issue 25) pp:
Publication Date(Web):May 27, 2010
DOI:10.1021/bi100571r
Arsenic is a carcinogen that tops the Superfund list of hazardous chemicals. Bacterial resistance to arsenic is facilitated by ArsD, which delivers As(III) to the ArsA ATPase, the catalytic subunit of the ArsAB pump. Here we report the structure of the arsenic metallochaperone ArsD at 1.4 Å and a model for its binding of metalloid. There are two ArsD molecules in the asymmetric unit. The overall structure of the ArsD monomer has a thioredoxin fold, with a core of four β-strands flanked by four α-helices. Based on data from structural homologues, ArsD was modeled with and without bound As(III). ArsD binds one arsenic per monomer coordinated with the three sulfur atoms of Cys12, Cys13, and Cys18. Using this structural model, an algorithm was used to dock ArsD and ArsA. The resulting docking model provides testable predictions of the contact points of the two proteins and forms the basis for future experiments.
Co-reporter:Chungang Yuan;Corinne R. Lehr;Jie Qin;Timothy R. McDermott;X. Chris Le
PNAS 2009 Volume 106 (Issue 13 ) pp:5213-5217
Publication Date(Web):2009-03-31
DOI:10.1073/pnas.0900238106
Arsenic is the most common toxic substance in the environment, ranking first on the Superfund list of hazardous substances.
It is introduced primarily from geochemical sources and is acted on biologically, creating an arsenic biogeocycle. Geothermal
environments are known for their elevated arsenic content and thus provide an excellent setting in which to study microbial
redox transformations of arsenic. To date, most studies of microbial communities in geothermal environments have focused on
Bacteria and Archaea, with little attention to eukaryotic microorganisms. Here, we show the potential of an extremophilic eukaryotic alga of the
order Cyanidiales to influence arsenic cycling at elevated temperatures. Cyanidioschyzon sp. isolate 5508 oxidized arsenite [As(III)] to arsenate [As(V)], reduced As(V) to As(III), and methylated As(III) to form
trimethylarsine oxide (TMAO) and dimethylarsenate [DMAs(V)]. Two arsenic methyltransferase genes, CmarsM7 and CmarsM8, were cloned from this organism and demonstrated to confer resistance to As(III) in an arsenite hypersensitive strain of
Escherichia coli. The 2 recombinant CmArsMs were purified and shown to transform As(III) into monomethylarsenite, DMAs(V), TMAO, and trimethylarsine
gas, with a Topt of 60–70 °C. These studies illustrate the importance of eukaryotic microorganisms to the biogeochemical cycling of arsenic
in geothermal systems, offer a molecular explanation for how these algae tolerate arsenic in their environment, and provide
the characterization of algal methyltransferases.
Co-reporter:Jiaojiao Li, Goutam Mandal, Barry P. Rosen
Anaerobe (June 2016) Volume 39() pp:117-123
Publication Date(Web):June 2016
DOI:10.1016/j.anaerobe.2016.03.012
Co-reporter:Hung-Chi Yang, Barry P. Rosen
Biomedical Journal (February 2016) Volume 39(Issue 1) pp:5-13
Publication Date(Web):February 2016
DOI:10.1016/j.bj.2015.08.003
Co-reporter:Venkadesh Sarkarai Nadar, Masafumi Yoshinaga, Shashank S. Pawitwar, Palani Kandavelu, ... Barry P. Rosen
Journal of Molecular Biology (5 June 2016) Volume 428(Issue 11) pp:2462-2473
Publication Date(Web):5 June 2016
DOI:10.1016/j.jmb.2016.04.022
•Arsenic is the most pervasive environmental toxin.•ArsI is a microbial non-heme, ferrous-dependent dioxygenase that detoxifies organoarsenical herbicides and growth promoters.•The structure of ArsI shows a novel organoarsenical binding site.•A loop-gating mechanism is proposed for substrate access to the catalytic site.Arsenic is a ubiquitous and carcinogenic environmental element that enters the biosphere primarily from geochemical sources, but also through anthropogenic activities. Microorganisms play an important role in the arsenic biogeochemical cycle by biotransformation of inorganic arsenic into organic arsenicals and vice versa. ArsI is a microbial non-heme, ferrous-dependent dioxygenase that transforms toxic methylarsenite [MAs(III)] to less toxic and carcinogenic inorganic arsenite [As(III)] by C–As bond cleavage. An ArsI ortholog, TcArsI, from the thermophilic bacterium Thermomonospora curvata was expressed, purified, and crystallized. The structure was solved in both the apo form and with Ni(II), Co(II), or Fe(III). The MAs(III) binding site is a vicinal cysteine pair in a flexible loop. A structure with the loop occupied with β-mercaptoethanol mimics binding of MAs(III). The structure of a mutant protein (Y100H/V102F) was solved in two different crystal forms with two other orientations of the flexible loop. These results suggest that a loop-gating mechanism controls the catalytic reaction. In the ligand-free open state, the loop is exposed to solvent, where it can bind MAs(III). The loop moves toward the active site, where it forms a closed state that orients the C–As bond for dioxygen addition and cleavage. Elucidation of the enzymatic mechanism of this unprecedented C–As lyase reaction will enhance our understanding of recycling of environmental organoarsenicals.Download high-res image (133KB)Download full-size image
.png)