Co-reporter:József Kalmár ; Kelemu L. Woldegiorgis ; Bernadett Biri
Journal of the American Chemical Society 2011 Volume 133(Issue 49) pp:19911-19921
Publication Date(Web):October 27, 2011
DOI:10.1021/ja2083152
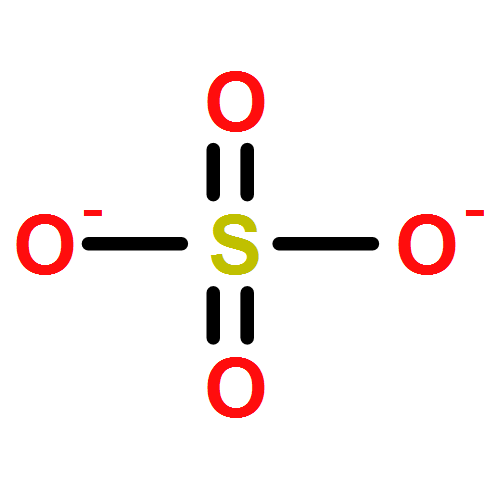
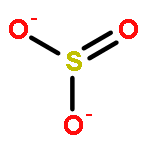
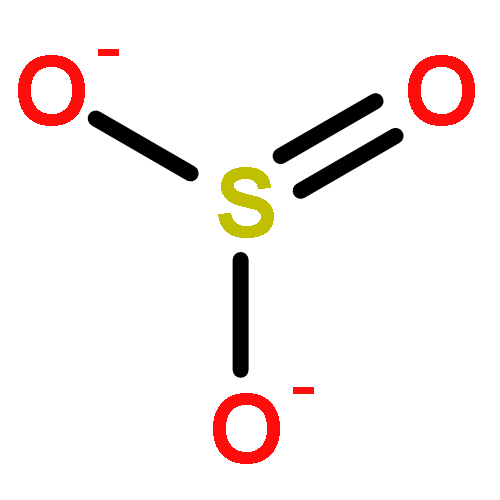
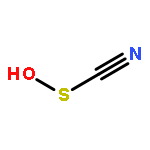
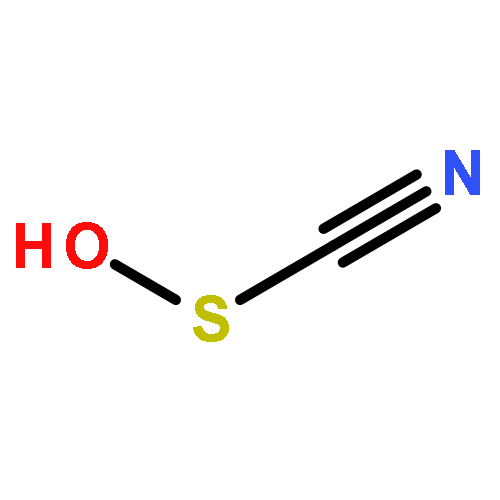
Relatively little is known about the reaction chemistry of the human defense factor hypothiocyanite (OSCN–) and its conjugate acid hypothiocyanous acid (HOSCN), in part because of their instability in aqueous solutions. Herein we report that HOSCN/OSCN– can engage in a cascade of pH- and concentration-dependent comproportionation, disproportionation, and hydrolysis reactions that control its stability in water. On the basis of reaction kinetic, spectroscopic, and chromatographic methods, a detailed mechanism is proposed for the decomposition of HOSCN/OSCN– in the range of pH 4–7 to eventually give simple inorganic anions including CN–, OCN–, SCN–, SO32–, and SO42–. Thiocyanogen ((SCN)2) is proposed to be a key intermediate in the hydrolysis; and the facile reaction of (SCN)2 with OSCN– to give NCS(═O)SCN, a previously unknown reactive sulfur species, has been independently investigated. The mechanism of the aqueous decomposition of (SCN)2 around pH 4 is also reported. The resulting mechanistic models for the decomposition of HOSCN and (SCN)2 address previous empirical observations, including the facts that the presence of SCN– and/or (SCN)2 decreases the stability of HOSCN/OSCN–, that radioisotopic labeling provided evidence that under physiological conditions decomposing OSCN– is not in equilibrium with (SCN)2 and SCN–, and that the hydrolysis of (SCN)2 near neutral pH does not produce OSCN–. Accordingly, we demonstrate that, during the human peroxidase-catalyzed oxidation of SCN–, (SCN)2 cannot be the precursor of the OSCN– that is produced.
Co-reporter:Bheki A. Xulu and Michael T. Ashby
Biochemistry 2010 Volume 49(Issue 9) pp:
Publication Date(Web):January 19, 2010
DOI:10.1021/bi902089w
Thiocyanate reacts noncatalytically with myeloperoxidase-derived HOCl to produce hypothiocyanite (OSCN−), thereby potentially limiting the propensity of HOCl to inflict host tissue damage that can lead to inflammatory diseases. However, the efficiency with which SCN− captures HOCl in vivo depends on the concentration of SCN− relative to other chemical targets. In blood plasma, where the concentration of SCN− is relatively low, proteins may be the principal initial targets of HOCl, and chloramines are a significant product. Chloramines eventually decompose to irreversibly damage proteins. In the present study, we demonstrate that SCN− reacts efficiently with chloramines in small molecules, in proteins, and in Escherichia coli cells to give OSCN− and the parent amine. Remarkably, OSCN− reacts faster than SCN− with chloramines. These reactions of SCN− and OSCN− with chloramines may repair some of the damage that is inflicted on protein amines by HOCl. Our observations are further evidence for the importance of secondary reactions during the redox cascades that are associated with oxidative stress by hypohalous acids.
Co-reporter:Hisanori Ueki, Garry Chapman and Michael T. Ashby
The Journal of Physical Chemistry A 2010 Volume 114(Issue 4) pp:1670-1676
Publication Date(Web):December 29, 2009
DOI:10.1021/jp906651n
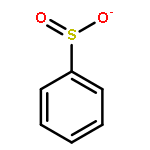
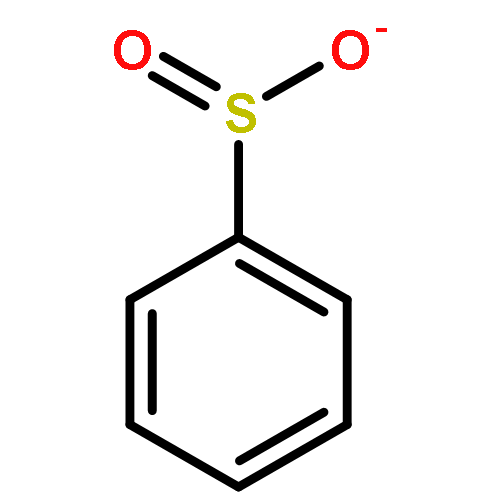
The mechanism of oxidation of ArSO2− (PhSO2− and 5-sulfinato-2-nitrobenzoic acid = TNBO21−/2−) with HOCl/OCl− has been investigated using the kinetic method. In contrast to previous reports for PhSO2− (for which it was suggested that OCl− and not HOCl was the reactant), the reaction proceeds through a conventional pathway: nucleophilic attack by ArSO2− on HOCl with concomitant Cl+ transfer to give a sulfonyl chloride intermediate (ArSO2Cl), which we have identified spectrophotometrically. Remarkably, the rate constant for the reaction of HOCl with ArSO2− is on the order of 109 M−1 s−1, larger than the rate constants for corresponding thiolates, and is nearly diffusion-controlled. In contrast, the rate constant for the reaction of OCl− with ArSO2− is approximately 7 orders of magnitude smaller.
Co-reporter:Kelemu Lemma and Michael T. Ashby
Chemical Research in Toxicology 2009 Volume 22(Issue 9) pp:1622
Publication Date(Web):August 25, 2009
DOI:10.1021/tx900212r
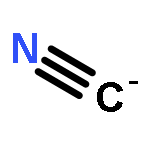
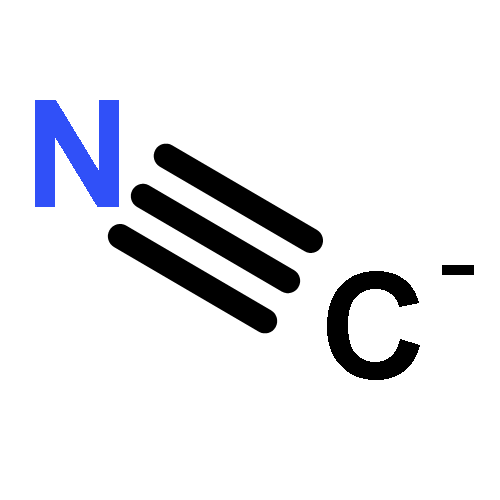
The chief sources of cyanide (CN−) in humans are tobacco and occupationally derived smoke, inflammation [vis-à-vis myeloperoxidase (MPO)-induced chlorination of glycine], and microbial cyanogenesis (including Pseudomonas aeruginosa infection of the cystic fibrosis lung). The human mucosae of healthy individuals are usually protected from infection by innate defense mechanisms that include the defensive peroxidase systems. In the oral cavity, salivary peroxidase and MPO catalyze the oxidation of the pseudohalide thiocyanate (SCN−) by hydrogen peroxide to produce the antimicrobial hypothiocyanite (OSCN−). Lactoperoxidase carries out the same reaction in the human lung (as does MPO during inflammatory response). In the present study, we show that OSCN− and CN− react with pH-dependent kinetics to produce SCN− and cyanate (OCN−) via dicyanosulfide (NCSCN), with the maximum rate occurring near neutral, physiological pH. In addition to presenting a detailed chemical mechanism, we discuss unresolved issues, including the possible biological relevance of the NCSCN intermediate.
Co-reporter:Jennifer L. Beal, Steven B. Foster and Michael T. Ashby
Biochemistry 2009 Volume 48(Issue 46) pp:
Publication Date(Web):October 19, 2009
DOI:10.1021/bi901343d
Electrophilic halogenating agents, including hypohalous acids and haloamines, oxidize free methionine and the N-terminal methionines of peptides and proteins (e.g., Met-1 of anti-inflammatory peptide 1 and ubiquitin) to produce dehydromethionine (a five-membered isothiazolidinium heterocycle). Amide derivatives of methionine are oxidized to the corresponding sulfoxide derivatives under the same reaction conditions (e.g., Met-3 of anti-inflammatory peptide 1). Other biological oxidants, including hydrogen peroxide and peroxynitrite, also produce only the corresponding sulfoxides. Hypothiocyanite does not react with methionine residues. We suggest that dehydromethionine may be a useful biomarker for the myeloperoxidase-induced oxidative stress associated with many inflammatory diseases.
Co-reporter:D. Tim Harwood, Susan L. Nimmo, Anthony J. Kettle, Christine C. Winterbourn and Michael T. Ashby
Chemical Research in Toxicology 2008 Volume 21(Issue 5) pp:1011
Publication Date(Web):May 1, 2008
DOI:10.1021/tx800050n
Reduced glutathione (GSH) is a cornerstone of the antioxidant stratagem for eukaryotes and some prokaryotes. Hypochlorous acid (HOCl), which is produced by neutrophilic myeloperoxidase, reacts rapidly with excess GSH to yield mainly oxidized glutathione (GSSG). GSSG can be further oxidized to give first N-chloro derivatives and, later, higher oxidation states at the S centers. Under certain conditions, another major species that is observed during the oxidation of GSH by HOCl (and a minor species for other oxidants) exhibits a molecular mass that is 30 mass units heavier than GSH. This GSH+2O-2H species, which has been employed as a biomarker for oxidative stress, has been previously proposed to be a sulfonamide. Employing NMR spectroscopy and mass spectrometry, we demonstrate that the GSH+2O-2H species is indeed a nine-membered cyclic sulfonamide. Alternative formulations, including six-membered 1,2,5-oxathiazine heterocycles, have been ruled out. Remarkably, the sulfonamide exists as a 2:1 equilibrium mixture of two diastereomers. Isotope tracer studies have demonstrated that it is the Glu Cα center that has undergone racemization. It is proposed that the racemization takes place via an acyclic imine−sulfinic acid intermediate. The glutathione sulfonamides are stable products of GSH that have been detected in physiological systems. Elucidation of the structures of the glutathione sulfonamides provides further impetus to explore their potential as biomarkers of hypochlorous acid formation.
Co-reporter:Xiaoguang Wang and Michael T. Ashby
Chemical Research in Toxicology 2008 Volume 21(Issue 11) pp:2120
Publication Date(Web):October 22, 2008
DOI:10.1021/tx800195n
Hypothiocyanite (OSCN−) is a putative antimicrobial that is produced by defensive human peroxidases, including salivary peroxidase, lactoperoxidase, eosinophil peroxidase, and myeloperoxidase. The reaction of OSCN− with cysteine-derived sulfhydryl groups is believed to be involved in the antimicrobial mechanism of action. Hypothiocyanite decomposes via an unknown mechanism that involves multiple redox and hydrolytic steps to eventually yield mixtures of CN−/OCN− and SO32−/SO42−. Until recently, no information was available regarding the chemical nature of the intermediate(s) that are produced during the chemical cascade, but we have shown that OSCN− undergoes hydrolysis to give the new compound thiocarbamate-S-oxide, H2NC(═O)SO−. In the present paper, we demonstrate that H2NC(═O)SO− reacts with cysteine (CySH) via a two-step mechanism that is analogous to that of OSCN− with cysteine: HOZ + CySH → CySZ + H2O and CySZ + CySH → CySSCy + HZ, where Z = SCN and SC(═O)NH2, respectively. The kinetics and mechanism of both steps of the reaction of H2NC(═O)SO− with CySH have been investigated as a function of pH.
Co-reporter:Péter Nagy and Michael T. Ashby
Chemical Research in Toxicology 2007 Volume 20(Issue 9) pp:1364
Publication Date(Web):September 1, 2007
DOI:10.1021/tx700168z
The kinetics and mechanisms of the hydrolysis of cysteine thiosulfinate ester (CyS(═O)SCyx−, x = 0–2) have been investigated by stopped-flow spectrophotometry between pH 6 and pH 14. The rate-limiting reaction of hydroxide is observed for pH < 13. More complicated kinetics are observed above pH 13, where the hydrolysis of CyS(═O)SCy2− can be fast relative to subsequent reactions. The eventual products of hydrolysis are a 1:1 molar ratio of cystine (CySSCy) and cysteine sufinic acid (CySO2H) under all reaction conditions. The rate of hydrolysis is dependent upon the proton state of CyS(═O)SCyx−. Furthermore, cysteine thiosulfonate ester (CyS(═O)2SCy) was observed as an intermediate during the hydrolysis of CyS(═O)SCyx− at lower pH. CyS(═O)2SCy eventually hydrolyzes to give stoichiometric amounts of CySSCy and CySO2H. However, CySO2H is observed under some conditions for which hydrolysis of CyS(═O)2SCy is relatively slow, thus suggesting multiple hydrolysis pathways for CyS(═O)SCyx−. The mechanism up to the rate-limiting step is proposed to be as follows: CyS(═O)SCy0 = H+ + CyS(═O)SCy−, pKa3 = 7.32; CyS(═O)SCy− = H+ + CyS(═O)SCy2−, pKa4 = 7.92; CyS(═O)SCy0 + OH− → products, P0k0 = (5.0 ± 0.01) × 103 M−1 s−1; CyS(═O)SCy− + OH− → products, P1k1 = 60 ± 18 M−1 s−1; and CyS(═O)SCy2− + OH− → products, P2k2 = 0.36 ± 0.01 M−1 s−1, where Px is a constant (1 ≤ Px ≤ 3) that accounts for the partitioning between the possible hydrolysis pathways and the stoichiometries of their net reactions.
Co-reporter:Kelemu Lemma;Susan L. Alguindigue Nimmo
Heteroatom Chemistry 2007 Volume 18(Issue 5) pp:467-471
Publication Date(Web):10 JUL 2007
DOI:10.1002/hc.20340
Cysteine sulfenyl thiocyanate (CSSCN) reacts with thiols at pH 0 to cleanly yield disulfides. 2-Mercaptoethanol (2-MESH), 3-mercaptopropionic acid (3-MPASH), penicillamine (PENSH), and glutathione (GSH) react with CSSCN to give the corresponding mixed disulfides: 2-MESSC, 3-MPASSC, PENSSC, and GSSC. These compounds are stable at pH 0 and have been characterized by 1H and 13C NMR spectroscopy. © 2007 Wiley Periodicals, Inc. 18:467–471, 2007; Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/hc.20340
Co-reporter:Michael T. Ashby;Péter Nagy
Journal of Pharmaceutical Sciences 2006 Volume 95(Issue 1) pp:15-18
Publication Date(Web):18 NOV 2005
DOI:10.1002/jps.20521
Co-reporter:Vincent Bonifay, Tessa J. Barrett, David I. Pattison, Michael J. Davies, Clare L. Hawkins, Michael T. Ashby
Archives of Biochemistry and Biophysics (15 December 2014) Volume 564() pp:
Publication Date(Web):15 December 2014
DOI:10.1016/j.abb.2014.08.014
•Thiocyanate-derived oxidants induce Trp oxidation at pH values below 2.•A series of mono- and di-oxygenated Trp products are formed.•Evidence is presented for generation of Trp species with m/z 320.1, 336.1 and 261.1.•Similar products are formed on exposure of proteins to HOCl and HOBr.•Oxidized Trp residues are not suitable biomarkers to assess HOSCN in vivo.Human defensive peroxidases, including lactoperoxidase (LPO) and myeloperoxidase (MPO), are capable of catalyzing the oxidation of halides (X−) by H2O2 to give hypohalous acids (HOX) for the purpose of cellular defense. Substrate selectivity depends upon the relative abundance of the halides, but the pseudo-halide thiocyanate (SCN−) is a major substrate, and sometimes the exclusive substrate, of all defensive peroxidases in most physiologic fluids. The resulting hypothiocyanous acid (HOSCN) has been implicated in cellular damage via thiol oxidation. While thiols are believed to be the primary target of HOSCN in vivo, Trp residues have also been implicated as targets for HOSCN. However, the mechanism involved in HOSCN-mediated Trp oxidation was not established. Trp residues in proteins appeared to be susceptible to oxidation by HOSCN, whereas free Trp and Trp residues in small peptides were found to be unreactive. We show that HOSCN-induced Trp oxidation is dependent on pH, with oxidation of free Trp, and Trp-containing peptides observed when the pH is below 2. These conditions mimic those employed previously to precipitate proteins after treatment with HOSCN, which accounts for the discrepancy in the results reported for proteins versus free Trp and small peptides. The reactant in these cases may be thiocyanogen ((SCN)2), which is produced by comproportionation of HOSCN and SCN− at low pH. Reaction of thiocyanate-derived oxidants with protein Trp residues at low pH results in the formation of a number of oxidation products, including mono- and di-oxygenated derivatives, which are also formed with other hypohalous acids. Our data suggest that significant modification of Trp by HOSCN in vivo is likely to have limited biological relevance.
.png)