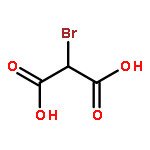
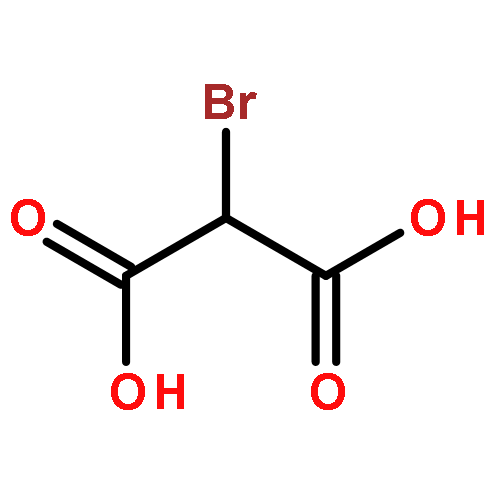
Co-reporter:Bruce Rodenborn;Chih-Hung Chen;Bin Liu;H. P. Zhang
PNAS 2013 110 (5 ) pp:
Publication Date(Web):2013-01-29
DOI:10.1073/pnas.1219831110
The swimming of a bacterium or a biomimetic nanobot driven by a rotating helical flagellum is often interpreted using the
resistive force theory developed by Gray and Hancock and by Lighthill, but this theory has not been tested for a range of
physically relevant parameters. We test resistive force theory in experiments on macroscopic swimmers in a fluid that is highly
viscous so the Reynolds number is small compared to unity, just as for swimming microorganisms. The measurements are made
for the range of helical wavelengths λ, radii R, and lengths L relevant to bacterial flagella. The experiments determine thrust, torque, and drag, thus providing a complete description
of swimming driven by a rotating helix at low Reynolds number. Complementary numerical simulations are conducted using the
resistive force theories, the slender body theories of Lighthill and Johnson, and the regularized Stokeslet method. The experimental
results differ qualitatively and quantitatively from the predictions of resistive force theory. The difference is especially
large for and/or , parameter ranges common for bacteria. In contrast, the predictions of Stokeslet and slender body analyses agree with the
laboratory measurements within the experimental uncertainty (a few percent) for all λ, R, and L. We present code implementing the slender body, regularized Stokeslet, and resistive force theories; thus readers can readily
compute force, torque, and drag for any bacterium or nanobot driven by a rotating helical flagellum.
Co-reporter:Bruce Rodenborn;Chih-Hung Chen;Bin Liu;H. P. Zhang
PNAS 2013 110 (5 ) pp:
Publication Date(Web):2013-01-29
DOI:10.1073/pnas.1219831110
The swimming of a bacterium or a biomimetic nanobot driven by a rotating helical flagellum is often interpreted using the
resistive force theory developed by Gray and Hancock and by Lighthill, but this theory has not been tested for a range of
physically relevant parameters. We test resistive force theory in experiments on macroscopic swimmers in a fluid that is highly
viscous so the Reynolds number is small compared to unity, just as for swimming microorganisms. The measurements are made
for the range of helical wavelengths λ, radii R, and lengths L relevant to bacterial flagella. The experiments determine thrust, torque, and drag, thus providing a complete description
of swimming driven by a rotating helix at low Reynolds number. Complementary numerical simulations are conducted using the
resistive force theories, the slender body theories of Lighthill and Johnson, and the regularized Stokeslet method. The experimental
results differ qualitatively and quantitatively from the predictions of resistive force theory. The difference is especially
large for and/or , parameter ranges common for bacteria. In contrast, the predictions of Stokeslet and slender body analyses agree with the
laboratory measurements within the experimental uncertainty (a few percent) for all λ, R, and L. We present code implementing the slender body, regularized Stokeslet, and resistive force theories; thus readers can readily
compute force, torque, and drag for any bacterium or nanobot driven by a rotating helical flagellum.
Co-reporter:H. P. Zhang;Avraham Be’er;E.-L. Florin;
Proceedings of the National Academy of Sciences 2010 107(31) pp:13626-13630
Publication Date(Web):July 19, 2010
DOI:10.1073/pnas.1001651107
Flocking birds, fish schools, and insect swarms are familiar examples of collective motion that plays a role in a range of
problems, such as spreading of diseases. Models have provided a qualitative understanding of the collective motion, but progress
has been hindered by the lack of detailed experimental data. Here we report simultaneous measurements of the positions, velocities,
and orientations as a function of time for up to a thousand wild-type Bacillus subtilis bacteria in a colony. The bacteria spontaneously form closely packed dynamic clusters within which they move cooperatively.
The number of bacteria in a cluster exhibits a power-law distribution truncated by an exponential tail. The probability of
finding clusters with large numbers of bacteria grows markedly as the bacterial density increases. The number of bacteria
per unit area exhibits fluctuations far larger than those for populations in thermal equilibrium. Such “giant number fluctuations”
have been found in models and in experiments on inert systems but not observed previously in a biological system. Our results
demonstrate that bacteria are an excellent system to study the general phenomenon of collective motion.
Co-reporter:Avraham Be’er;Gil Ariel;Oren Kalisman;Yael Helman;Alexandra Sirota-Madi;H.P. Zhang;E.-L. Florin;Shelley M. Payne;Eshel Ben-Jacob;
Proceedings of the National Academy of Sciences 2010 107(14) pp:6258-6263
Publication Date(Web):March 22, 2010
DOI:10.1073/pnas.1001062107
Sibling Paenibacillus dendritiformis bacterial colonies grown on low-nutrient agar medium mutually inhibit growth through secretion of a lethal factor. Analysis
of secretions reveals the presence of subtilisin (a protease) and a 12 kDa protein, termed sibling lethal factor (Slf). Purified
subtilisin promotes the growth and expansion of P. dendritiformis colonies, whereas Slf is lethal and lyses P. dendritiformis cells in culture. Slf is encoded by a gene belonging to a large family of bacterial genes of unknown function, and the gene
is predicted to encode a protein of approximately 20 kDa, termed dendritiformis sibling bacteriocin. The 20 kDa recombinant protein was produced and found to be inactive, but exposure to subtilisin resulted
in cleavage to the active, 12 kDa form. The experimental results, combined with mathematical modeling, show that subtilisin
serves to regulate growth of the colony. Below a threshold concentration, subtilisin promotes colony growth and expansion.
However, once it exceeds a threshold, as occurs at the interface between competing colonies, Slf is then secreted into the
medium to rapidly reduce cell density by lysis of the bacterial cells. The presence of genes encoding homologs of dendritiformis sibling bacteriocin in other bacterial species suggests that this mechanism for self-regulation of colony growth might not
be limited to P. dendritiformis.
Co-reporter:Avraham Be'er;H. P. Zhang;E.-L. Florin;Shelley M. Payne;Eshel Ben-Jacob
PNAS 2009 Volume 106 (Issue 2 ) pp:428-433
Publication Date(Web):2009-01-13
DOI:10.1073/pnas.0811816106
Bacteria can secrete a wide array of antibacterial compounds when competing with other bacteria for the same resources. Some
of these compounds, such as bacteriocins, can affect bacteria of similar or closely related strains. In some cases, these
secretions have been found to kill sibling cells that belong to the same colony. Here, we present experimental observations
of competition between 2 sibling colonies of Paenibacillus dendritiformis grown on a low-nutrient agar gel. We find that neighboring colonies (growing from droplet inoculation) mutually inhibit growth
through secretions that become lethal if the level exceeds a well-defined threshold. In contrast, within a single colony developing
from a droplet inoculation, no growth inhibition is observed. However, growth inhibition and cell death are observed if material
extracted from the agar between 2 growing colonies is introduced outside a growing single colony. To interpret the observations,
we devised a simple mathematical model for the secretion of an antibacterial compound. Simulations of this model illustrate
how secretions from neighboring colonies can be deadly, whereas secretions from a single colony growing from a droplet are
not.
Co-reporter:Robert S. Shaw;Norman Packard;Matthias Schröter;
Proceedings of the National Academy of Sciences 2007 104(23) pp:9580-9584
Publication Date(Web):May 23, 2007
DOI:10.1073/pnas.0703280104
Past work has shown that ions can pass through a membrane more readily in one direction than the other. We demonstrate here
in a model and an experiment that for a mixture of small and large particles such asymmetric diffusion can arise solely from
an asymmetry in the geometry of the pores of the membrane. Our deterministic simulation considers a two-dimensional gas of
elastic disks of two sizes diffusing through a membrane, and our laboratory experiment examines the diffusion of glass beads
of two sizes through a metal membrane. In both experiment and simulation, the membrane is permeable only to the smaller particles,
and the asymmetric pores lead to an asymmetry in the diffusion rates of these particles. The presence of even a small percentage
of large particles can clog a membrane, preventing passage of the small particles in one direction while permitting free flow
of the small particles in the other direction. The purely geometric kinetic constraints may play a role in common biological
contexts such as membrane ion channels.
.jpg)