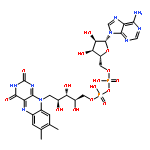
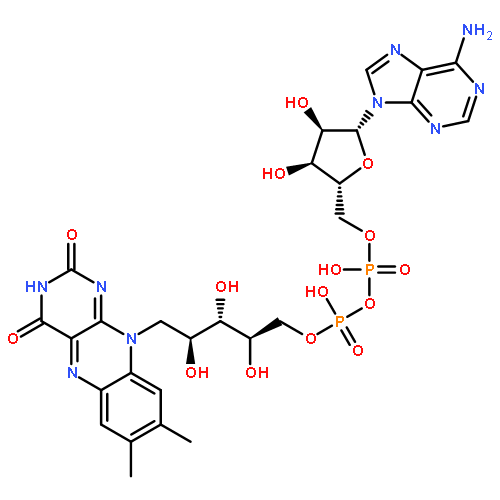
Co-reporter:Amie K. Boal and Amy C. Rosenzweig
Journal of the American Chemical Society October 14, 2009 Volume 131(Issue 40) pp:14196-14197
Publication Date(Web):September 16, 2009
DOI:10.1021/ja906363t
Copper trafficking proteins, including the chaperone Atox1 and the P1B-type ATPase ATP7B, have been implicated in cellular resistance to the anticancer drug cisplatin. We have determined two crystal structures of cisplatin-Atox1 adducts that reveal platinum coordination by the conserved CXXC copper-binding motif. Direct interaction of cisplatin with this functionally relevant site has significant implications for understanding the molecular basis for resistance mediated by copper transport pathways.
Co-reporter:Laura M. K. Dassama;Grace E. Kenney
Metallomics (2009-Present) 2017 vol. 9(Issue 1) pp:7-20
Publication Date(Web):2017/01/26
DOI:10.1039/C6MT00208K
Methanobactins (Mbns) are ribosomally produced, post-translationally modified peptide (RiPP) natural products that bind copper with high affinity using nitrogen-containing heterocycles and thioamide groups. In some methanotrophic bacteria, Mbns are secreted under conditions of copper starvation and then re-internalized as a copper source for the enzyme particulate methane monooxygenase (pMMO). Genome mining studies have led to the identification and classification of operons encoding the Mbn precursor peptide (MbnA) as well as a number of putative transport, regulatory, and biosynthetic proteins. These Mbn operons are present in non-methanotrophic bacteria as well, suggesting a broader role in and perhaps beyond copper acquisition. Genetic and biochemical studies indicate that specific operon-encoded proteins are involved in Mbn transport and provide insight into copper-responsive gene regulation in methanotrophs. Mbn biosynthesis is not yet understood, but combined analysis of Mbn structures, MbnA sequences, and operon content represents a powerful approach to elucidating the roles of specific biosynthetic enzymes. Future work will likely lead to the discovery of unique pathways for natural product biosynthesis and new mechanisms of microbial metal homeostasis.
Co-reporter:Matthew O. Ross
JBIC Journal of Biological Inorganic Chemistry 2017 Volume 22( Issue 2-3) pp:
Publication Date(Web):
DOI:10.1007/s00775-016-1419-y
Co-reporter:Aaron T. Smith, Matthew O. Ross, Brian M. HoffmanAmy C. Rosenzweig
Biochemistry 2017 Volume 56(Issue 1) pp:
Publication Date(Web):December 8, 2016
DOI:10.1021/acs.biochem.6b01022
The P1B-ATPases, a family of transmembrane metal transporters important for transition metal homeostasis in all organisms, are subdivided into classes based on sequence conservation and metal specificity. The multifunctional P1B-4-ATPase CzcP is part of the cobalt, zinc, and cadmium resistance system from the metal-tolerant, model organism Cupriavidus metallidurans. Previous work revealed the presence of an unusual soluble metal-binding domain (MBD) at the CzcP N-terminus, but the nature, extent, and selectivity of the transmembrane metal-binding site (MBS) of CzcP have not been resolved. Using homology modeling, we show that four wholly conserved amino acids from the transmembrane (TM) domain (Met254, Ser474, Cys476, and His807) are logical candidates for the TM MBS, which may communicate with the MBD via interactions with the first TM helix. Metal-binding analyses indicate that wild-type (WT) CzcP has three MBSs, and data on N-terminally truncated (ΔMBD) CzcP suggest the presence of a single TM MBS. Electronic absorption and electron paramagnetic resonance spectroscopic analyses of ΔMBD CzcP and variant proteins thereof provide insight into the details of Co2+ coordination by the TM MBS. These spectroscopic data, combined with in vitro functional studies of WT and variant CzcP proteins, show that the side chains of Met254, Cys476, and His807 contribute to Cd2+, Co2+, and Zn2+ binding and transport, whereas the side chain of Ser474 appears to play a minimal role. By comparison to other P1B-4-ATPases, we suggest that an evolutionarily adapted flexibility in the TM region likely afforded CzcP the ability to transport Cd2+ and Zn2+ in addition to Co2+.
Co-reporter:Grace E. Kenney, Anthony W. Goering, Matthew O. Ross, Caroline J. DeHart, Paul M. Thomas, Brian M. Hoffman, Neil L. Kelleher, and Amy C. Rosenzweig
Journal of the American Chemical Society 2016 Volume 138(Issue 35) pp:11124-11127
Publication Date(Web):August 15, 2016
DOI:10.1021/jacs.6b06821
Methanobactins (Mbns) are a growing family of ribosomally produced, post-translationally modified natural products. Characteristic nitrogen-containing heterocycles and neighboring thioamides allow these compounds to bind copper with high affinity. Genome mining has enabled the identification of Mbn operons in bacterial genomes and the prediction of diverse Mbn structures from operon content and precursor peptide sequence. Here we report the characterization of Mbn from Methylosinus (Ms.) species (sp.) LW4. The peptide backbone is distinct from all previously characterized Mbns, and the post-translational modifications correspond precisely to those predicted on the basis of the Ms. sp. LW4 Mbn operon. Thus, prediction based on genome analysis combined with isolation and structural characterization represents a phylogenetic approach to finding diverse Mbns and elucidating their biosynthetic pathways.
Co-reporter:Thomas J. Lawton
Journal of the American Chemical Society 2016 Volume 138(Issue 30) pp:9327-9340
Publication Date(Web):July 1, 2016
DOI:10.1021/jacs.6b04568
Biological conversion of natural gas to liquids (Bio-GTL) represents an immense economic opportunity. In nature, aerobic methanotrophic bacteria and anaerobic archaea are able to selectively oxidize methane using methane monooxygenase (MMO) and methyl coenzyme M reductase (MCR) enzymes. Although significant progress has been made toward genetically manipulating these organisms for biotechnological applications, the enzymes themselves are slow, complex, and not recombinantly tractable in traditional industrial hosts. With turnover numbers of 0.16–13 s–1, these enzymes pose a considerable upstream problem in the biological production of fuels or chemicals from methane. Methane oxidation enzymes will need to be engineered to be faster to enable high volumetric productivities; however, efforts to do so and to engineer simpler enzymes have been minimally successful. Moreover, known methane-oxidizing enzymes have different expression levels, carbon and energy efficiencies, require auxiliary systems for biosynthesis and function, and vary considerably in terms of complexity and reductant requirements. The pros and cons of using each methane-oxidizing enzyme for Bio-GTL are considered in detail. The future for these enzymes is bright, but a renewed focus on studying them will be critical to the successful development of biological processes that utilize methane as a feedstock.
Co-reporter:Thomas J. Lawton, Grace E. Kenney, Joseph D. Hurley, and Amy C. Rosenzweig
Biochemistry 2016 Volume 55(Issue 15) pp:2278-2290
Publication Date(Web):March 24, 2016
DOI:10.1021/acs.biochem.6b00175
The CopC proteins are periplasmic copper binding proteins believed to play a role in bacterial copper homeostasis. Previous studies have focused on CopCs that are part of seven-protein Cop or Pco systems involved in copper resistance. These canonical CopCs contain distinct Cu(I) and Cu(II) binding sites. Mounting evidence suggests that CopCs are more widely distributed, often present only with the CopD inner membrane protein, frequently as a fusion protein, and that the CopC and CopD proteins together function in the uptake of copper to the cytoplasm. In the methanotroph Methylosinus trichosporium OB3b, genes encoding a CopCD pair are located adjacent to the particulate methane monooxygenase (pMMO) operon. The CopC from this organism (Mst-CopC) was expressed, purified, and structurally characterized. The 1.46 Å resolution crystal structure of Mst-CopC reveals a single Cu(II) binding site with coordination somewhat different from that in canonical CopCs, and the absence of a Cu(I) binding site. Extensive bioinformatic analyses indicate that the majority of CopCs in fact contain only a Cu(II) site, with just 10% of sequences corresponding to the canonical two-site CopC. Accordingly, a new classification scheme for CopCs was developed, and detailed analyses of the sequences and their genomic neighborhoods reveal new proteins potentially involved in copper homeostasis, providing a framework for expanded models of CopCD function.
Co-reporter:Laura M. K. Dassama;Grace E. Kenney;Soo Y. Ro;Eliza L. Zielazinski
PNAS 2016 Volume 113 (Issue 46 ) pp:13027-13032
Publication Date(Web):2016-11-15
DOI:10.1073/pnas.1603578113
Methanotrophic bacteria use methane, a potent greenhouse gas, as their primary source of carbon and energy. The first step
in methane metabolism is its oxidation to methanol. In almost all methanotrophs, this chemically challenging reaction is catalyzed
by particulate methane monooxygenase (pMMO), a copper-dependent integral membrane enzyme. Methanotrophs acquire copper (Cu)
for pMMO by secreting a small ribosomally produced, posttranslationally modified natural product called methanobactin (Mbn).
Mbn chelates Cu with high affinity, and the Cu-loaded form (CuMbn) is reinternalized into the cell via an active transport
process. Bioinformatic and gene regulation studies suggest that two proteins might play a role in CuMbn handling: the TonB-dependent
transporter MbnT and the periplasmic binding protein MbnE. Disruption of the gene that encodes MbnT abolishes CuMbn uptake,
as reported previously, and expression of MbnT in Escherichia coli confers the ability to take up CuMbn. Biophysical studies of MbnT and MbnE reveal specific interactions with CuMbn, and a
crystal structure of apo MbnE is consistent with MbnE's proposed role as a periplasmic CuMbn transporter. Notably, MbnT and
MbnE exhibit different levels of discrimination between cognate and noncognate CuMbns. These findings provide evidence for
CuMbn–protein interactions and begin to elucidate the molecular mechanisms of its recognition and transport.
Co-reporter:Sarah Sirajuddin and Amy C. Rosenzweig
Biochemistry 2015 Volume 54(Issue 14) pp:2283-2294
Publication Date(Web):March 25, 2015
DOI:10.1021/acs.biochem.5b00198
Methane monooxygenases (MMOs) are enzymes that catalyze the oxidation of methane to methanol in methanotrophic bacteria. As potential targets for new gas-to-liquid methane bioconversion processes, MMOs have attracted intense attention in recent years. There are two distinct types of MMO, a soluble, cytoplasmic MMO (sMMO) and a membrane-bound, particulate MMO (pMMO). Both oxidize methane at metal centers within a complex, multisubunit scaffold, but the structures, active sites, and chemical mechanisms are completely different. This Current Topic review article focuses on the overall architectures, active site structures, substrate reactivities, protein–protein interactions, and chemical mechanisms of both MMOs, with an emphasis on fundamental aspects. In addition, recent advances, including new details of interactions between the sMMO components, characterization of sMMO intermediates, and progress toward understanding the pMMO metal centers are highlighted. The work summarized here provides a guide for those interested in exploiting MMOs for biotechnological applications.
Co-reporter:Megen A. Culpepper ; George E. Cutsail III ; William A. Gunderson ; Brian M. Hoffman
Journal of the American Chemical Society 2014 Volume 136(Issue 33) pp:11767-11775
Publication Date(Web):July 24, 2014
DOI:10.1021/ja5053126
Particulate methane monooxygenase (pMMO) catalyzes the oxidation of methane to methanol in methanotrophic bacteria. As a copper-containing enzyme, pMMO has been investigated extensively by electron paramagnetic resonance (EPR) spectroscopy, but the presence of multiple copper centers has precluded correlation of EPR signals with the crystallographically identified monocopper and dicopper centers. A soluble recombinant fragment of the pmoB subunit of pMMO, spmoB, like pMMO itself, contains two distinct copper centers and exhibits methane oxidation activity. The spmoB protein, spmoB variants designed to disrupt one or the other or both copper centers, as well as native pMMO have been investigated by EPR, ENDOR, and ESEEM spectroscopies in combination with metal content analysis. The data are remarkably similar for spmoB and pMMO, validating the use of spmoB as a model system. The results indicate that one EPR-active Cu(II) ion is present per pMMO and that it is associated with the active-site dicopper center in the form of a valence localized Cu(I)Cu(II) pair; the Cu(II), however, is scrambled between the two locations within the dicopper site. The monocopper site observed in the crystal structures of pMMO can be assigned as Cu(I). 14N ENDOR and ESEEM data are most consistent with one of these dicopper-site signals involving coordination of the Cu(II) ion by residues His137 and His139, the other with Cu(II) coordinated by His33 and the N-terminal amino group. 1H ENDOR measurements indicate there is no aqua (HxO) ligand bound to the Cu(II), either terminally or as a bridge to Cu(I).
Co-reporter:Megen A. Culpepper and Amy C. Rosenzweig
Biochemistry 2014 Volume 53(Issue 39) pp:6211-6219
Publication Date(Web):September 3, 2014
DOI:10.1021/bi500850j
In the initial steps of their metabolic pathway, methanotrophic bacteria oxidize methane to methanol with methane monooxygenases (MMOs) and methanol to formaldehyde with methanol dehydrogenases (MDHs). Several lines of evidence suggest that the membrane-bound or particulate MMO (pMMO) and MDH interact to form a metabolic supercomplex. To further investigate the possible existence of such a supercomplex, native MDH from Methylococcus capsulatus (Bath) has been purified and characterized by size exclusion chromatography with multi-angle light scattering and X-ray crystallography. M. capsulatus (Bath) MDH is primarily a dimer in solution, although an oligomeric species with a molecular mass of ∼450–560 kDa forms at higher protein concentrations. The 2.57 Å resolution crystal structure reveals an overall fold and α2β2 dimeric architecture similar to those of other MDH structures. In addition, biolayer interferometry studies demonstrate specific protein–protein interactions between MDH and M. capsulatus (Bath) pMMO as well as between MDH and the truncated recombinant periplasmic domains of M. capsulatus (Bath) pMMO (spmoB). These interactions exhibit KD values of 833 ± 409 nM and 9.0 ± 7.7 μM, respectively. The biochemical data combined with analysis of the crystal lattice interactions observed in the MDH structure suggest a model in which MDH and pMMO associate not as a discrete, stoichiometric complex but as a larger assembly scaffolded by the intracytoplasmic membranes.
Co-reporter:Aaron T. Smith;Kyle P. Smith
JBIC Journal of Biological Inorganic Chemistry 2014 Volume 19( Issue 6) pp:947-960
Publication Date(Web):2014 August
DOI:10.1007/s00775-014-1129-2
The P1B-ATPases are integral membrane proteins that couple ATP hydrolysis to metal cation transport. Widely distributed across all domains of life, these enzymes have been previously shown to transport copper, zinc, cobalt, and other thiophilic heavy metals. Recent data suggest that these enzymes may also be involved in nickel and/or iron transport. Here we have exploited large amounts of genomic data to examine and classify the various P1B-ATPase subfamilies. Specifically, we have combined new methods of data partitioning and network visualization known as Transitivity Clustering and Protein Similarity Networks with existing biochemical data to examine properties such as length, speciation, and metal-binding motifs of the P1B-ATPase subfamily sequences. These data reveal interesting relationships among the enzyme sequences of previously established subfamilies, indicate the presence of two new subfamilies, and suggest the existence of new regulatory elements in certain subfamilies. Taken together, these findings underscore the importance of P1B-ATPases in homeostasis of nearly every biologically relevant transition metal and provide an updated framework for future studies.
Co-reporter:Eliza L. Zielazinski, Manuel González-Guerrero, Poorna Subramanian, Timothy L. Stemmler, José M. Argüello and Amy C. Rosenzweig
Metallomics 2013 vol. 5(Issue 12) pp:1614-1623
Publication Date(Web):03 Sep 2013
DOI:10.1039/C3MT00195D
The P1B-ATPases are a ubiquitous family of metal transporters. These transporters are classified into subfamilies on the basis of substrate specificity, which is conferred by conserved amino acids in the last three transmembrane domains. Five subfamilies have been identified to date, and representative members of four (P1B-1 to P1B-4) have been studied. The fifth family (P1B-5), of which some members contain a C-terminal hemerythrin (Hr) domain, is less well characterized. The S. meliloti Sma1163 gene encodes for a P1B-5-ATPase, denoted Nia (Nickel–iron ATPase), that is induced by exogenous Fe2+ and Ni2+. The nia mutant accumulates nickel and iron, suggesting a possible role in detoxification of these two elements under free-living conditions, as well as in symbiosis, when the highest expression levels are measured. This function is supported by an inhibitory effect of Fe2+ and Ni2+ on the pNPPase activity, and by the ability of Nia to bind Fe2+ in the transmembrane domain. Optical and X-ray absorption spectroscopic studies of the isolated Hr domain confirm the presence of a dinuclear iron center and suggest that this domain might function as an iron sensor.
Co-reporter:Laura M. K. Dassama ; Amie K. Boal ; Carsten Krebs ; Amy C. Rosenzweig ;J. Martin Bollinger ; Jr.
Journal of the American Chemical Society 2012 Volume 134(Issue 5) pp:2520-2523
Publication Date(Web):January 12, 2012
DOI:10.1021/ja211314p
The reaction of a class I ribonucleotide reductase (RNR) begins when a cofactor in the β subunit oxidizes a cysteine residue ∼35 Å away in the α subunit, generating a thiyl radical. In the class Ic enzyme from Chlamydia trachomatis (Ct), the cysteine oxidant is the MnIV ion of a MnIV/FeIII cluster, which assembles in a reaction between O2 and the MnII/FeII complex of β. The heterodinuclear nature of the cofactor raises the question of which site, 1 or 2, contains the MnIV ion. Because site 1 is closer to the conserved location of the cysteine-oxidizing tyrosyl radical of class Ia and Ib RNRs, we suggested that the MnIV ion most likely resides in this site (i.e., 1MnIV/2FeIII), but a subsequent computational study favored its occupation of site 2 (1FeIII/2MnIV). In this work, we have sought to resolve the location of the MnIV ion in Ct RNR-β by correlating X-ray crystallographic anomalous scattering intensities with catalytic activity for samples of the protein reconstituted in vitro by two different procedures. In samples containing primarily MnIV/FeIII clusters, Mn preferentially occupies site 1, but some anomalous scattering from site 2 is observed, implying that both 1MnII/2FeII and 1FeII/2MnII complexes are competent to react with O2 to produce the corresponding oxidized states. However, with diminished MnII loading in the reconstitution, there is no evidence for Mn occupancy of site 2, and the greater activity of these “low-Mn” samples on a per-Mn basis implies that the 1MnIV/2FeIII-β is at least the more active of the two oxidized forms and may be the only active form.
Co-reporter:Megen A. Culpepper, George E. Cutsail, III, Brian M. Hoffman, and Amy C. Rosenzweig
Journal of the American Chemical Society 2012 Volume 134(Issue 18) pp:7640-7643
Publication Date(Web):April 27, 2012
DOI:10.1021/ja302195p
Particulate methane monooxygenase (pMMO) is an integral membrane metalloenzyme that converts methane to methanol in methanotrophic bacteria. The enzyme consists of three subunits, pmoB, pmoA, and pmoC, organized in an α3β3γ3 trimer. Studies of intact pMMO and a recombinant soluble fragment of the pmoB subunit (denoted as spmoB) indicate that the active site is located within the soluble region of pmoB at the site of a crystallographically modeled dicopper center. In this work, we have investigated the reactivity of pMMO and spmoB with oxidants. Upon reduction and treatment of spmoB with O2 or H2O2 or pMMO with H2O2, an absorbance feature at 345 nm is generated. The energy and intensity of this band are similar to those of the μ-η2:η2-peroxo-CuII2 species formed in several dicopper enzymes and model compounds. The feature is not observed in inactive spmoB variants in which the dicopper center is disrupted, consistent with O2 binding to the proposed active site. Reaction of the 345 nm species with CH4 results in the disappearance of the spectroscopic feature, suggesting that this O2 intermediate is mechanistically relevant. Taken together, these observations provide strong new support for the identity and location of the pMMO active site.
Co-reporter:Grace E. Kenney and Amy C. Rosenzweig
ACS Chemical Biology 2012 Volume 7(Issue 2) pp:260
Publication Date(Web):November 29, 2011
DOI:10.1021/cb2003913
Methanotrophic bacteria, organisms that oxidize methane, produce a small copper chelating molecule called methanobactin (Mb). Mb binds Cu(I) with high affinity and is hypothesized to mediate copper acquisition from the environment. Recent advances in Mb characterization include revision of the chemical structure of Mb from Methylosinus trichosporium OB3b and further investigation of its biophysical properties. In addition, Mb production by several other methanotroph strains has been investigated, and preliminary characterization suggests diversity in chemical composition. Initial clues into Mb biosynthesis have been obtained by identification of a putative precursor gene in the M. trichosporium OB3b genome. Finally, direct uptake of intact Mb into the cytoplasm of M. trichosporium OB3b cells has been demonstrated, and studies of the transport mechanism have been initiated. Taken together, these advances represent significant progress and set the stage for exciting new research directions.
Co-reporter:Eliza L. Zielazinski, George E. Cutsail III, Brian M. Hoffman, Timothy L. Stemmler, and Amy C. Rosenzweig
Biochemistry 2012 Volume 51(Issue 40) pp:
Publication Date(Web):September 12, 2012
DOI:10.1021/bi3006708
The P1B-type ATPases are a ubiquitous family of P-type ATPases involved in the transport of transition metal ions. Divided into subclasses based on sequence characteristics and substrate specificity, these integral membrane transporters play key roles in metal homeostasis, metal tolerance, and the biosynthesis of metalloproteins. The P1B-4-ATPases have the simplest architecture of the five P1B-ATPase families and have been suggested to play a role in Co2+ transport. A P1B-4-ATPase from Sulfitobacter sp. NAS-14.1, designated sCoaT, has been cloned, expressed, and purified. Activity assays indicate that sCoaT is specific for Co2+. A single Co2+ binding site is present, and optical, electron paramagnetic resonance, and X-ray absorption spectroscopic data are consistent with tetrahedral coordination by oxygen and nitrogen ligands, including a histidine and likely a water. Surprisingly, there is no evidence for coordination by sulfur. Mutation of a conserved cysteine residue, Cys 327, in the signature transmembrane Ser-Pro-Cys metal binding motif does not abolish the ATP hydrolysis activity or affect the spectroscopic analysis, establishing that this residue is not involved in the initial Co2+ binding by sCoaT. In contrast, replacements of conserved transmembrane residues Ser 325, His 657, Glu 658, and Thr 661 with alanine abolish ATP hydrolysis activity and Co2+ binding, indicating that these residues are necessary for Co2+ transport. These data represent the first in vitro characterization of a P1B-4-ATPase and its Co2+ binding site.
Co-reporter:Amie K. Boal, Joseph A. Cotruvo Jr., JoAnne Stubbe, and Amy C. Rosenzweig
Biochemistry 2012 Volume 51(Issue 18) pp:3861-3871
Publication Date(Web):March 23, 2012
DOI:10.1021/bi201925t
Class Ib ribonucleotide reductases (RNRs) use a dimanganese-tyrosyl radical cofactor, MnIII2-Y•, in their homodimeric NrdF (β2) subunit to initiate reduction of ribonucleotides to deoxyribonucleotides. The structure of the MnII2 form of NrdF is an important component in understanding O2-mediated formation of the active metallocofactor, a subject of much interest because a unique flavodoxin, NrdI, is required for cofactor assembly. Biochemical studies and sequence alignments suggest that NrdF and NrdI proteins diverge into three phylogenetically distinct groups. The only crystal structure to date of a NrdF with a fully ordered and occupied dimanganese site is that of Escherichia coli MnII2-NrdF, prototypical of the enzymes from actinobacteria and proteobacteria. Here we report the 1.9 Å resolution crystal structure of Bacillus subtilis MnII2-NrdF, representative of the enzymes from a second group, from Bacillus and Staphylococcus. The structures of the metal clusters in the β2 dimer are distinct from those observed in E. coli MnII2-NrdF. These differences illustrate the key role that solvent molecules and protein residues in the second coordination sphere of the MnII2 cluster play in determining conformations of carboxylate residues at the metal sites and demonstrate that diverse coordination geometries are capable of serving as starting points for MnIII2-Y• cofactor assembly in class Ib RNRs.
Co-reporter:Stephen M. Smith, Swati Rawat, Joshua Telser, Brian M. Hoffman, Timothy L. Stemmler, and Amy C. Rosenzweig
Biochemistry 2011 Volume 50(Issue 47) pp:
Publication Date(Web):October 20, 2011
DOI:10.1021/bi200801z
Particulate methane monooxygenase (pMMO) is an integral membrane metalloenzyme that oxidizes methane to methanol in methanotrophic bacteria. Previous biochemical and structural studies of pMMO have focused on preparations from Methylococcus capsulatus (Bath) and Methylosinus trichosporium OB3b. A pMMO from a third organism, Methylocystis species strain M, has been isolated and characterized. Both membrane-bound and solubilized Methylocystis sp. strain M pMMO contain ∼2 copper ions per 100 kDa protomer and exhibit copper-dependent propylene epoxidation activity. Spectroscopic data indicate that Methylocystis sp. strain M pMMO contains a mixture of CuI and CuII, of which the latter exhibits two distinct type 2 CuII electron paramagnetic resonance (EPR) signals. Extended X-ray absorption fine structure (EXAFS) data are best fit with a mixture of Cu–O/N and Cu–Cu ligand environments with a Cu–Cu interaction at 2.52–2.64 Å. The crystal structure of Methylocystis sp. strain M pMMO was determined to 2.68 Å resolution and is the best quality pMMO structure obtained to date. It provides a revised model for the pmoA and pmoC subunits and has led to an improved model of M. capsulatus (Bath) pMMO. In these new structures, the intramembrane zinc/copper binding site has a different coordination environment from that in previous models.
Co-reporter:Neela H. Yennawar;Monica I. McLaughlin;Squire J. Booker;Amie K. Boal;Tyler L. Grove
Science 2011 Volume 332(Issue 6033) pp:1089-1092
Publication Date(Web):27 May 2011
DOI:10.1126/science.1205358
An enzyme harnesses methyl donation and reductive cleavage of its cofactor within a single active site to methylate RNA.
Co-reporter:Matthew E. Traverso, Poorna Subramanian, Roman Davydov, Brian M. Hoffman, Timothy L. Stemmler and Amy C. Rosenzweig
Biochemistry 2010 Volume 49(Issue 33) pp:
Publication Date(Web):July 23, 2010
DOI:10.1021/bi100866b
The P1B-type ATPases couple the energy of ATP hydrolysis to metal ion translocation across cell membranes. Important for prokaryotic metal resistance and essential metal distribution in eukaryotes, P1B-ATPases are divided into subclasses on the basis of their metal substrate specificities. Sequence analysis of putative P1B-5-ATPases, for which the substrate has not been identified, led to the discovery of a C-terminal soluble domain homologous to hemerythrin (Hr) proteins and domains. The Hr domain from the Acidothermus cellulolyticus P1B-5-ATPase was cloned, expressed, and purified (P1B-5-Hr). P1B-5-Hr binds two iron ions per monomer and adopts a predominantly helical fold. Optical absorption features of the iron-loaded and azide-treated protein are consistent with features observed for other Hr proteins. Autoxidation to the met form is very rapid, as reported for other prokaryotic Hr domains. The presence of a diiron center was confirmed by electron paramagnetic resonance (EPR) and X-ray absorption spectroscopic (XAS) data. The occurrence of a Hr-like domain in a P-type ATPase is unprecedented and suggests new regulatory mechanisms as well as an expanded function for Hr proteins in biology.
Co-reporter:Uchechi E. Ukaegbu, Auric Kantz, Michelle Beaton, George T. Gassner and Amy C. Rosenzweig
Biochemistry 2010 Volume 49(Issue 8) pp:
Publication Date(Web):January 7, 2010
DOI:10.1021/bi901693u
Styrene monooxygenase (SMO) is a two-component flavoprotein monooxygenase that transforms styrene to styrene oxide in the first step of the styrene catabolic and detoxification pathway of Pseudomonas putida S12. The crystal structure of the N-terminally histidine-tagged epoxidase component of this system, NSMOA, determined to 2.3 Å resolution, indicates the enzyme exists as a homodimer in which each monomer forms two distinct domains. The overall architecture is most similar to that of p-hydroxybenzoate hydroxylase (PHBH), although there are some significant differences in secondary structure. Structural comparisons suggest that a large cavity open to the surface forms the FAD binding site. At the base of this pocket is another cavity that likely represents the styrene binding site. Flavin binding and redox equilibria are tightly coupled such that reduced FAD binds apo NSMOA ∼8000 times more tightly than the oxidized coenzyme. Equilibrium fluorescence and isothermal titration calorimetry data using benzene as a substrate analogue indicate that the oxidized flavin and substrate analogue binding equilibria of NSMOA are linked such that the binding affinity of each is increased by 60-fold when the enzyme is saturated with the other. A much weaker ∼2-fold positive cooperative interaction is observed for the linked binding equilibria of benzene and reduced FAD. The low affinity of the substrate analogue for the reduced FAD complex of NSMOA is consistent with a preferred reaction order in which flavin reduction and reaction with oxygen precede the binding of styrene, identifying the apoenzyme structure as the key catalytic resting state of NSMOA poised to bind reduced FAD and initiate the oxygen reaction.
Co-reporter:Amie K. Boal and Amy C. Rosenzweig
Chemical Reviews 2009 Volume 109(Issue 10) pp:4760
Publication Date(Web):July 15, 2009
DOI:10.1021/cr900104z
Co-reporter:Uchechi E. Ukaegbu and Amy C. Rosenzweig
Biochemistry 2009 Volume 48(Issue 10) pp:
Publication Date(Web):February 20, 2009
DOI:10.1021/bi8019614
MmoS from Methylococcus capsulatus (Bath) is the multidomain sensor protein of a two-component signaling system proposed to play a role in the copper-mediated regulation of soluble methane monooxygenase (sMMO). MmoS binds an FAD cofactor within its N-terminal tandem Per-Arnt-Sim (PAS) domains, suggesting that it functions as a redox sensor. The crystal structure of the MmoS tandem PAS domains, designated PAS-A and PAS-B, has been determined to 2.34 Å resolution. Both domains adopt the typical PAS domain α/β topology and are structurally similar. The two domains are linked by a long α helix and do not interact with one another. The FAD cofactor is housed solely within PAS-A and is stabilized by an extended hydrogen bonding network. The overall fold of PAS-A is similar to those of other flavin-containing PAS domains, but homodimeric interactions in other structures are not observed in the MmoS sensor, which crystallized as a monomer. The structure both provides new insight into the architecture of tandem PAS domains and suggests specific residues that may play a role in MmoS FAD redox chemistry and subsequent signal transduction.
Co-reporter:Amanda S. Hakemian, Kalyan C. Kondapalli, Joshua Telser, Brian M. Hoffman, Timothy L. Stemmler and Amy C. Rosenzweig
Biochemistry 2008 Volume 47(Issue 26) pp:
Publication Date(Web):June 10, 2008
DOI:10.1021/bi800598h
Particulate methane monooxygenase (pMMO) is a membrane-bound metalloenzyme that oxidizes methane to methanol in methanotrophic bacteria. The nature of the pMMO active site and the overall metal content are controversial, with spectroscopic and crystallographic data suggesting the presence of a mononuclear copper center, a dinuclear copper center, a trinuclear center, and a diiron center or combinations thereof. Most studies have focused on pMMO from Methylococcus capsulatus (Bath). pMMO from a second organism, Methylosinus trichosporium OB3b, has been purified and characterized by spectroscopic and crystallographic methods. Purified M. trichosporium OB3b pMMO contains ∼2 copper ions per 100 kDa protomer. Electron paramagnetic resonance (EPR) spectroscopic parameters indicate that type 2 Cu(II) is present as two distinct species. Extended X-ray absorption fine structure (EXAFS) data are best fit with oxygen/nitrogen ligands and reveal a Cu−Cu interaction at 2.52 Å. Correspondingly, X-ray crystallography of M. trichosporium OB3b pMMO shows a dinuclear copper center, similar to that observed previously in the crystal structure of M. capsulatus (Bath) pMMO. There are, however, significant differences between the pMMO structures from the two organisms. A mononuclear copper center present in M. capsulatus (Bath) pMMO is absent in M. trichosporium OB3b pMMO, whereas a metal center occupied by zinc in the M. capsulatus (Bath) pMMO structure is occupied by copper in M. trichosporium OB3b pMMO. These findings extend previous work on pMMO from M. capsulatus (Bath) and provide new insight into the functional importance of the different metal centers.
Co-reporter:Liliya A. Yatsunyk;J. Allen Easton
JBIC Journal of Biological Inorganic Chemistry 2008 Volume 13( Issue 2) pp:271-288
Publication Date(Web):2008 February
DOI:10.1007/s00775-007-0320-0
ZnuA is the periplasmic Zn2+-binding protein associated with the high-affinity ATP-binding cassette ZnuABC transporter from Escherichia coli. Although several structures of ZnuA and its homologs have been determined, details regarding metal ion stoichiometry, affinity, and specificity as well as the mechanism of metal uptake and transfer remain unclear. The crystal structures of E. coli ZnuA (Eco-ZnuA) in the apo, Zn2+-bound, and Co2+-bound forms have been determined. ZnZnuA binds at least two metal ions. The first, observed previously in other structures, is coordinated tetrahedrally by Glu59, His60, His143, and His207. Replacement of Zn2+ with Co2+ results in almost identical coordination geometry at this site. The second metal binding site involves His224 and several yet to be identified residues from the His-rich loop that is unique to Zn2+ periplasmic metal binding receptors. Electron paramagnetic resonance and X-ray absorption spectroscopic data on CoZnuA provide additional insight into possible residues involved in this second site. The second site is also detected by metal analysis and circular dichroism (CD) titrations. Eco-ZnuA binds Zn2+ (estimated Kd < 20 nM), Co2+, Ni2+, Cu2+, Cu+, and Cd2+, but not Mn2+. Finally, conformational changes upon metal binding observed in the crystal structures together with fluorescence and CD data indicate that only Zn2+ substantially stabilizes ZnuA and might facilitate recognition of ZnuB and subsequent metal transfer.
Co-reporter:Ramakrishnan Balasubramanian and Amy C. Rosenzweig
Accounts of Chemical Research 2007 Volume 40(Issue 7) pp:573
Publication Date(Web):April 20, 2007
DOI:10.1021/ar700004s
Particulate methane monooxygense (pMMO) is an integral membrane copper-containing enzyme that converts methane to methanol. Knowledge of how pMMO selectively oxidizes methane under ambient conditions could impact the development of new catalysts. The crystal structure of Methylococcus capsulatus (Bath) pMMO reveals the composition and location of three metal centers. Spectroscopic data provide insight into the coordination environments and oxidation states of these metal centers. These results, combined with computational studies and comparisons to relevant systems, are discussed in the context of identifying the most likely site for O2 activation.
Co-reporter:Raquel L. Lieberman and Amy C. Rosenzweig
Dalton Transactions 2005 (Issue 21) pp:3390-3396
Publication Date(Web):26 Sep 2005
DOI:10.1039/B506651D
Particulate methane monooxygenase is a copper-containing, membrane-bound metalloenzyme that converts methane to methanol in Nature. How pMMO accomplishes this difficult reaction under ambient conditions is one of the major unsolved problems in bioinorganic chemistry. Despite considerable research efforts in the past 20 years, the active site of the enzyme remains unknown. We recently solved the first crystal structure of pMMO to 2.8 Å resolution, revealing the overall structure, oligomerization state, subunit ratio, and composition and location of the metal centers. Almost none of the key structural features were predicted. In this Perspective, we review the state of knowledge before and after the structure determination, emphasizing elucidation of the pMMO active site.
Co-reporter:Raquel L. Lieberman
and
Amy C. Rosenzweig
Nature 2005 434(7030) pp:177
Publication Date(Web):
DOI:10.1038/nature03311
Co-reporter:Raquel L. Lieberman;Deepak B. Shrestha;Peter E. Doan;Brian M. Hoffman;Timothy L. Stemmler;
Proceedings of the National Academy of Sciences 2003 100(7) pp:3820-3825
Publication Date(Web):March 12, 2003
DOI:10.1073/pnas.0536703100
Particulate methane monooxygenase (pMMO) is a membrane-bound enzyme that catalyzes the oxidation of methane to methanol in
methanotropic bacteria. Understanding how this enzyme hydroxylates methane at ambient temperature and pressure is of fundamental
chemical and potential commercial importance. Difficulties in solubilizing and purifying active pMMO have led to conflicting
reports regarding its biochemical and biophysical properties, however. We have purified pMMO from Methylococcus capsulatus (Bath) and detected activity. The purified enzyme has a molecular mass of ≈200 kDa, probably corresponding to an α2β2γ2 polypeptide arrangement. Each 200-kDa pMMO complex contains 4.8 ± 0.8 copper ions and 1.5 ± 0.7 iron ions. Electron paramagnetic
resonance spectroscopic parameters corresponding to 40–60% of the total copper are consistent with the presence of a mononuclear
type 2 copper site. X-ray absorption near edge spectra indicate that purified pMMO is a mixture of Cu(I) and Cu(II) oxidation
states. Finally, extended x-ray absorption fine structure data are best fit with oxygen/nitrogen ligands and a 2.57-Å Cu-Cu
interaction, providing direct evidence for a copper-containing cluster in pMMO.
Co-reporter:
Nature Structural and Molecular Biology 2001 8(9) pp:751 - 755
Publication Date(Web):
DOI:10.1038/nsb0901-751
Co-reporter:Walter C. Voegtli;Jie Ge;Deborah L. Perlstein;JoAnne Stubbe
PNAS 2001 Volume 98 (Issue 18 ) pp:10073-10078
Publication Date(Web):2001-08-28
DOI:10.1073/pnas.181336398
The R2 subunits of class I ribonucleotide reductases (RNRs) house a diferric-tyrosyl radical (Y·) cofactor essential for DNA
synthesis. In yeast, there are two R2 proteins, Y2 and Y4. Although both Y2 and Y4 are homologous to R2s from other organisms,
Y4 lacks three conserved iron-binding residues, and its exact function is unclear. Y4 is required for assembly of the diferric-Y·
cofactor in Y2, and the two proteins can form both homodimeric and heterodimeric complexes. The Y2Y4 heterodimer was crystallized
from a mixture of the two proteins, and its structure was determined to 2.8 Å resolution. Both Y2 and Y4 are completely α
helical and resemble the mouse and Escherichia coli R2s in overall fold. Three α helices not observed in the mouse R2 structure are present at the Y2 N terminus, and one extra
N-terminal helix is observed in Y4. In addition, one of the eight principal helices in both Y2 and Y4, αD, is shifted significantly
from its position in mouse R2. The heterodimer interface is similar to the mouse R2 homodimer interface in size and interacting
residues, but loop regions at the interface edges differ. A single metal ion, assigned as Zn(II), occupies the Fe2 position
in the Y2 active site. Treatment of the crystals with Fe(II) results in difference electron density consistent with formation
of a diiron center. No metal-binding site is observed in Y4. Instead, the residues in the active site region form a hydrogen-bonding
network involving an arginine, two glutamic acids, and a water molecule.
Co-reporter:Matthew T. Miller;Brian O. Bachmann;Craig A. Townsend
Nature Structural and Molecular Biology 2001 8(8) pp:684-689
Publication Date(Web):2001-08-01
DOI:10.1038/90394
The enzyme -lactam synthetase (-LS) catalyzes the formation of the -lactam ring in clavulanic acid, a clinically important -lactamase inhibitor. Whereas the penicillin -lactam ring is generated by isopenicillin N synthase (IPNS) in the presence of ferrous ion and dioxygen, -LS uses ATP and Mg2+ as cofactors. According to sequence alignments, -LS is homologous to class B asparagine synthetases (AS-Bs), ATP/Mg2+-dependent enzymes that convert aspartic acid to asparagine. Here we report the first crystal structure of a -LS. The 1.95 Å resolution structure of Streptomyces clavuligerus -LS provides a fully resolved view of the active site in which substrate, closely related ATP analog ,-methyleneadenosine 5'-triphosphate (AMP-CPP) and a single Mg2+ ion are present. A high degree of substrate preorganization is observed. Comparison to Escherichia coli AS-B reveals the evolutionary changes that have taken place in -LS that impede interdomain reaction, which is essential in AS-B, and that accommodate -lactam formation. The structural data provide the opportunity to alter the synthetic potential of -LS, perhaps leading to the creation of new -lactamase inhibitors and -lactam antibiotics.
Co-reporter:
Nature Structural and Molecular Biology 2000 7(9) pp:766-771
Publication Date(Web):
DOI:10.1038/78999
The Hah1 metallochaperone protein is implicated in copper delivery to the Menkes and Wilson disease proteins. Hah1 and the N-termini of its target proteins belong to a family of metal binding domains characterized by a conserved MT/HCXXC sequence motif. The crystal structure of Hah1 has been determined in the presence of Cu(I), Hg(II), and Cd(II). The 1.8 Å resolution structure of CuHah1 reveals a copper ion coordinated by Cys residues from two adjacent Hah1 molecules. The CuHah1 crystal structure is the first of a copper chaperone bound to copper and provides structural support for direct metal ion exchange between conserved MT/HCXXC motifs in two domains. The structures of HgHah1 and CdHah1, determined to 1.75 Å resolution, also reveal metal ion coordination by two MT/HCXXC motifs. An extended hydrogen bonding network, unique to the complex of two Hah1 molecules, stabilizes the metal binding sites and suggests specific roles for several conserved residues. Taken together, the structures provide models for intermediates in metal ion transfer and suggest a detailed molecular mechanism for protein recognition and metal ion exchange between MT/HCXXC containing domains.
Co-reporter:Thomas J Lawton, Amy C Rosenzweig
Current Opinion in Chemical Biology (December 2016) Volume 35() pp:
Publication Date(Web):1 December 2016
DOI:10.1016/j.cbpa.2016.10.001
•The MCR reaction proceeds via a Ni(II)-thiolate intermediate.•A recombinant expression system for ANME MCR has been developed.•The structure of a key intermediate in the sMMO catalytic cycle has been elucidated.•The crystal structure of an sMMO protein–protein complex has been determined.•Catalytically active pMMO has been incorporated into a printable polymer.Nature utilizes two groups of enzymes to catalyze methane conversions, methyl-coenzyme M reductases (MCRs) and methane monooxygenases (MMOs). These enzymes have been difficult to incorporate into industrial processes due to their complexity, poor stability, and lack of recombinant tractability. Despite these issues, new ways of preparing and stabilizing these enzymes have recently been discovered, and new mechanistic insight into how MCRs and MMOs break the C–H bond in nature's most inert hydrocarbon have been obtained. This review focuses on recent findings in the methane biocatalysis field, and discusses the impact of these finding on designing MMO and MCR-based biotechnologies.Download high-res image (214KB)Download full-size image
Co-reporter:Monika Sommerhalter, Yongbo Zhang, Amy C. Rosenzweig
Journal of Molecular Biology (19 January 2007) Volume 365(Issue 3) pp:715-721
Publication Date(Web):19 January 2007
DOI:10.1016/j.jmb.2006.10.030
COMMD1 is the prototype of a new protein family that plays a role in several important cellular processes, including NF-κB signaling, sodium transport, and copper metabolism. The COMMD proteins interact with one another via a conserved C-terminal domain, whereas distinct functions are predicted to result from a variable N-terminal domain. The COMMD proteins have not been characterized biochemically or structurally. Here, we present the solution structure of the N-terminal domain of COMMD1 (N-COMMD1, residues 1–108). This domain adopts an α-helical structure that bears little resemblance to any other helical protein. The compact nature of N-COMMD1 suggests that full-length COMMD proteins are modular, consistent with specific functional properties for each domain. Interactions between N-COMMD1 and partner proteins may occur via complementary electrostatic surfaces. These data provide a new foundation for biochemical characterization of COMMD proteins and for probing COMMD1 protein–protein interactions at the molecular level.
Co-reporter:Amie K. Boal
Journal of the American Chemical Society () pp:
Publication Date(Web):September 16, 2009
DOI:10.1021/ja906363t
Copper trafficking proteins, including the chaperone Atox1 and the P1B-type ATPase ATP7B, have been implicated in cellular resistance to the anticancer drug cisplatin. We have determined two crystal structures of cisplatin-Atox1 adducts that reveal platinum coordination by the conserved CXXC copper-binding motif. Direct interaction of cisplatin with this functionally relevant site has significant implications for understanding the molecular basis for resistance mediated by copper transport pathways.
.jpg)














![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)